Cytologické úlohy na Jednotnú štátnu skúšku z biológie. Materiál na prípravu na Jednotnú štátnu skúšku (GIA) z biológie (11. ročník) na tému: Edukačná a metodická príručka Riešenie problémov z cytológie
Bunka ako biologický systém
Moderná bunková teória, jej hlavné ustanovenia, úloha pri formovaní moderného prírodovedného obrazu sveta. Rozvoj vedomostí o bunke. Bunková štruktúra organizmov je základom jednoty organického sveta, dôkazom príbuznosti živej prírody
Moderná bunková teória, jej hlavné ustanovenia, úloha pri formovaní moderného prírodovedného obrazu sveta
Jedným zo základných konceptov modernej biológie je myšlienka, že všetky živé organizmy majú bunkovú štruktúru. Veda študuje štruktúru bunky, jej životnú aktivitu a interakciu s prostredím. cytológie, teraz častejšie označovaný ako bunková biológia. Cytológia vďačí za svoj vznik formulácii bunkovej teórie (1838-1839, M. Schleiden, T. Schwann, doplnená v roku 1855 R. Virchowom).
Bunková teória je zovšeobecnená predstava o štruktúre a funkciách buniek ako živých jednotiek, ich reprodukcii a úlohe pri tvorbe mnohobunkových organizmov.
Základné princípy bunkovej teórie:
- Bunka je jednotka štruktúry, životnej aktivity, rastu a vývoja živých organizmov - mimo bunky neexistuje život.
- Bunka je jediný systém pozostávajúci z mnohých prvkov, ktoré sú navzájom prirodzene prepojené a predstavujú určitú integrálnu formáciu.
- Bunky všetkých organizmov sú podobné svojim chemickým zložením, štruktúrou a funkciami.
- Nové bunky vznikajú až delením materských buniek („bunka z bunky“).
- Bunky mnohobunkových organizmov tvoria tkanivá a orgány sa skladajú z tkanív. Život organizmu ako celku je určený interakciou buniek, ktoré ho tvoria.
- Bunky mnohobunkových organizmov majú celú sadu génov, líšia sa však od seba tým, že v nich pracujú rôzne skupiny génov, čoho výsledkom je morfologická a funkčná rôznorodosť buniek – diferenciácia.
Vďaka vytvoreniu bunkovej teórie sa ukázalo, že bunka je najmenšia jednotka života, elementárny živý systém, ktorý má všetky znaky a vlastnosti živých vecí. Formulácia bunkovej teórie sa stala najdôležitejším predpokladom pre rozvoj názorov na dedičnosť a variabilitu, pretože identifikácia ich povahy a inherentných vzorcov nevyhnutne naznačovala univerzálnosť štruktúry živých organizmov. Identifikácia jednoty chemického zloženia a štruktúry buniek slúžila ako impulz pre rozvoj predstáv o pôvode živých organizmov a ich vývoji. Vznik mnohobunkových organizmov z jednej bunky počas embryonálneho vývoja sa navyše stal dogmou modernej embryológie.
Rozvoj vedomostí o bunke
Až do 17. storočia ľudia nevedeli vôbec nič o mikroštruktúre predmetov okolo seba a svet vnímali voľným okom. Zariadenie na štúdium mikrosveta - mikroskop - vynašli okolo roku 1590 holandskí mechanici G. a Z. Jansenovci, no jeho nedokonalosť neumožňovala skúmať dostatočne malé predmety. K pokroku v tejto oblasti prispelo až vytvorenie tzv. zloženého mikroskopu K. Drebbela (1572-1634) na jeho základe.
V roku 1665 anglický fyzik R. Hooke (1635-1703) zdokonalil konštrukciu mikroskopu a technológiu brúsenia šošoviek a v snahe zabezpečiť lepšiu kvalitu obrazu skúmal pod ním časti korku, dreveného uhlia a živých rastlín. Na rezoch objavil drobné póry, pripomínajúce plást, a nazval ich bunky (z lat. celulum- bunka, bunka). Je zaujímavé, že R. Hooke považoval bunkovú membránu za hlavnú zložku bunky.
V druhej polovici 17. storočia sa objavili práce najvýznamnejších mikroskopistov M. Malpighiho (1628-1694) a N. Grewa (1641-1712), ktorí objavili aj bunkovú stavbu mnohých rastlín.
Aby sa uistil, že to, čo videli R. Hooke a ďalší vedci, je pravda, holandský obchodník A. van Leeuwenhoek, ktorý nemal žiadne špeciálne vzdelanie, nezávisle vyvinul dizajn mikroskopu, ktorý sa zásadne líšil od existujúceho, a zlepšil technológiu výroby šošoviek. To mu umožnilo dosiahnuť 275- až 300-násobné zväčšenie a preskúmať konštrukčné detaily, ktoré boli pre iných vedcov technicky nedostupné. A. van Leeuwenhoek bol neprekonateľný pozorovateľ: pozorne načrtol a opísal to, čo videl pod mikroskopom, no nesnažil sa to vysvetliť. Objavil jednobunkové organizmy vrátane baktérií a našiel jadrá, chloroplasty a zhrubnutie bunkových stien v rastlinných bunkách, no jeho objavy boli ocenené oveľa neskôr.
Objavy zložiek vnútornej stavby organizmov v prvej polovici 19. storočia nasledovali jeden za druhým. G. Mohl rozlíšil živú hmotu a vodnatú tekutinu - bunkovú šťavu - v rastlinných bunkách a objavil póry. Anglický botanik R. Brown (1773-1858) objavil jadro v bunkách orchideí v roku 1831, potom bolo objavené vo všetkých rastlinných bunkách. Český vedec J. Purkinje (1787-1869) vymyslel termín „protoplazma“ na označenie polotekutého želatínového obsahu bunky bez jadra (1840). Ďalej ako všetci jeho súčasníci pokročil belgický botanik M. Schleiden (1804-1881), ktorý štúdiom vývoja a diferenciácie rôznych bunkových štruktúr vyšších rastlín dokázal, že všetky rastlinné organizmy pochádzajú z jednej bunky. Skúmal tiež zaoblené telá jadierok v jadrách buniek cibuľových šupín (1842).
V roku 1827 ruský embryológ K. Baer objavil vajíčka ľudí a iných cicavcov, čím vyvrátil myšlienku vývoja organizmu výlučne z mužských gamét. Okrem toho dokázal vznik mnohobunkového živočíšneho organizmu z jedinej bunky – oplodneného vajíčka, ako aj podobnosť štádií embryonálneho vývoja mnohobunkových živočíchov, čo naznačovalo jednotu ich pôvodu. Informácie nahromadené do polovice 19. storočia si vyžadovali zovšeobecnenie, ktoré sa stalo bunkovou teóriou. Biológia vďačí za svoju formuláciu nemeckému zoológovi T. Schwannovi (1810-1882), ktorý na základe vlastných údajov a záverov M. Schleidena o vývoji rastlín vyslovil predpoklad, že ak je jadro prítomné v akomkoľvek útvare viditeľnom pod mikroskop, potom je tento útvar bunkový. Na základe tohto kritéria sformuloval T. Schwann hlavné ustanovenia bunkovej teórie.
Nemecký lekár a patológ R. Virchow (1821-1902) vniesol do tejto teórie ďalší dôležitý bod: bunky vznikajú len delením pôvodnej bunky, t. j. bunky vznikajú len z buniek („bunka z bunky“).
Od vzniku bunkovej teórie sa doktrína bunky ako jednotky štruktúry, funkcie a vývoja organizmu neustále rozvíja. Koncom 19. storočia sa vďaka úspechom mikroskopickej technológie podarilo objasniť štruktúru bunky, popísať organely – časti buniek, ktoré plnia rôzne funkcie, študovať spôsoby tvorby nových buniek (mitóza, meióza), a vyjasnil sa primárny význam bunkových štruktúr pri prenose dedičných vlastností. Použitie najnovších fyzikálno-chemických výskumných metód umožnilo hlbšie preniknúť do procesov ukladania a prenosu dedičných informácií, ako aj študovať jemnú štruktúru každej z bunkových štruktúr. To všetko prispelo k oddeleniu bunkovej vedy do samostatného odvetvia poznania - cytológie.
Bunková štruktúra organizmov, podobnosť štruktúry buniek všetkých organizmov je základom jednoty organického sveta, dôkazom príbuznosti živej prírody
Všetky dnes známe živé organizmy (rastliny, zvieratá, huby a baktérie) majú bunkovú štruktúru. Dokonca aj vírusy, ktoré nemajú bunkovú štruktúru, sa môžu množiť iba v bunkách. Bunka je základná stavebná a funkčná jednotka živej bytosti, ktorá sa vyznačuje všetkými svojimi prejavmi, najmä látkovou premenou a premenou energie, homeostázou, rastom a vývojom, rozmnožovaním a dráždivosťou. Zároveň sa v bunkách ukladajú, spracúvajú a implementujú dedičné informácie.
Napriek všetkej rozmanitosti buniek je štrukturálny plán pre ne rovnaký: všetky obsahujú dedičný aparátponorený do cytoplazme a okolitá bunka plazmatická membrána.
Bunka vznikla ako výsledok dlhého vývoja organického sveta. Spojenie buniek do mnohobunkového organizmu nie je jednoduchým súhrnom, pretože každá bunka, pričom si zachováva všetky vlastnosti, ktoré sú vlastné živému organizmu, zároveň získava nové vlastnosti vďaka svojej špecifickej funkcii. Na jednej strane možno mnohobunkový organizmus rozdeliť na jednotlivé časti - bunky, ale na druhej strane ich opätovným zložením nie je možné obnoviť funkcie celého organizmu, pretože iba v interakcii častí sa v systéme objavia nové vlastnosti. To odhaľuje jeden z hlavných vzorov, ktoré charakterizujú živé veci – jednotu diskrétneho a holistického. Malé veľkosti a značný počet buniek vytvárajú v mnohobunkových organizmoch veľký povrch potrebný na zabezpečenie rýchleho metabolizmu. Okrem toho, ak jedna časť tela odumrie, jej integrita sa môže obnoviť reprodukciou buniek. Mimo bunky je ukladanie a prenos dedičných informácií, ukladanie a prenos energie s jej následnou premenou na prácu nemožné. Napokon, rozdelenie funkcií medzi bunky v mnohobunkovom organizme poskytovalo organizmom dostatok príležitostí na prispôsobenie sa ich prostrediu a bolo predpokladom pre zvýšenie zložitosti ich organizácie.
Stanovenie jednoty štrukturálneho plánu buniek všetkých živých organizmov teda slúžilo ako dôkaz jednoty pôvodu všetkého života na Zemi.
Rozmanitosť buniek. Prokaryotické a eukaryotické bunky. Porovnávacie charakteristiky buniek rastlín, živočíchov, baktérií, húb Diverzita buniek
Bunka je podľa bunkovej teórie najmenšia štrukturálna a funkčná jednotka organizmov, ktorá má všetky vlastnosti živého tvora. Na základe počtu buniek sa organizmy delia na jednobunkové a mnohobunkové. Bunky jednobunkových organizmov existujú ako nezávislé organizmy a vykonávajú všetky funkcie živých vecí. Všetky prokaryoty a množstvo eukaryotov (veľa druhov rias, húb a prvokov) sú jednobunkové, ktoré udivujú mimoriadnou rozmanitosťou tvarov a veľkostí. Väčšina organizmov je však stále mnohobunková. Ich bunky sa špecializujú na vykonávanie určitých funkcií a tvoria tkanivá a orgány, ktoré môžu ovplyvniť ich morfologické vlastnosti. Napríklad ľudské telo je tvorené približne 1014 bunkami, ktoré predstavuje približne 200 druhov, ktoré majú širokú škálu tvarov a veľkostí.
Tvar buniek môže byť okrúhly, valcový, kubický, hranolový, diskovitý, vretenovitý, hviezdicovitý atď.. Vajíčka teda majú okrúhly tvar, epitelové bunky majú tvar valcový, kubický a hranolový, červené krvinky majú tvar bikonkávneho disku, bunky svalového tkaniva majú vretenovitý tvar a stelát - bunky nervového tkaniva. Množstvo buniek nemá vôbec žiadny stály tvar. Patria sem predovšetkým krvné leukocyty.
Veľkosti buniek sa tiež výrazne líšia: väčšina buniek mnohobunkového organizmu má veľkosti od 10 do 100 mikrónov a najmenšie - 2 až 4 mikróny. Spodná hranica je spôsobená skutočnosťou, že bunka musí mať minimálny súbor látok a štruktúr na zabezpečenie životne dôležitej činnosti a príliš veľká veľkosť bunky bude narúšať výmenu látok a energie s prostredím a bude tiež komplikovať procesy. udržiavania homeostázy. Niektoré bunky však možno vidieť aj voľným okom. V prvom rade ide o bunky melónu a jabĺk, ako aj vajíčka rýb a vtákov. Aj keď jeden z lineárnych rozmerov bunky presahuje priemer, všetky ostatné zodpovedajú norme. Napríklad proces neurónu môže presiahnuť dĺžku 1 m, ale jeho priemer bude stále zodpovedať priemernej hodnote. Neexistuje žiadny priamy vzťah medzi veľkosťou buniek a veľkosťou tela. Svalové bunky slona a myši sú teda rovnako veľké.
Prokaryotické a eukaryotické bunky
Ako bolo uvedené vyššie, bunky majú mnoho podobných funkčných vlastností a morfologických vlastností. Každá z nich pozostáva z cytoplazmy, ktorá je v nej ponorená dedičný aparát a oddelené od vonkajšieho prostredia plazmatická membrána, alebo plazmalema, ktorý nezasahuje do procesu metabolizmu a energie. Bunka môže mať mimo membrány aj bunkovú stenu pozostávajúcu z rôznych látok, ktorá slúži na ochranu bunky a je akousi vonkajšou kostrou.
Cytoplazma je celý obsah bunky, ktorý vypĺňa priestor medzi plazmatickou membránou a štruktúrou obsahujúcou genetickú informáciu. Pozostáva z hlavnej látky - hyaloplazma- a do nej ponorené organely a inklúzie. Organoidy sú trvalé zložky bunky, ktoré vykonávajú určité funkcie, a inklúzie sú zložky, ktoré sa objavujú a miznú počas života bunky, pričom primárne vykonávajú zásobné alebo vylučovacie funkcie. Inklúzie sú často rozdelené na pevné a kvapalné. Pevné inklúzie sú zastúpené hlavne granulami a môžu mať rôznu povahu, zatiaľ čo vakuoly a tukové kvapôčky sa považujú za tekuté inklúzie.

V súčasnosti existujú dva hlavné typy bunkovej organizácie: prokaryotické a eukaryotické.
Prokaryotická bunka nemá jadro, jej genetická informácia nie je oddelená od cytoplazmy membránami.
Oblasť cytoplazmy, v ktorej je uložená genetická informácia v prokaryotickej bunke, sa nazýva tzv nukleoid. V cytoplazme prokaryotických buniek sa vyskytuje hlavne jeden typ organel - ribozómy a organely obklopené membránami úplne chýbajú. Baktérie sú prokaryoty.
Eukaryotická bunka je bunka, v ktorej má aspoň jedno z vývojových štádií jadro- špeciálna štruktúra, v ktorej sa nachádza DNA.
Cytoplazma eukaryotických buniek sa vyznačuje významnou diverzitou membránových a nemembránových organel. Eukaryotické organizmy zahŕňajú rastliny, živočíchy a huby. Veľkosť prokaryotických buniek je zvyčajne rádovo menšia ako veľkosť eukaryotických buniek. Väčšina prokaryotov sú jednobunkové organizmy, zatiaľ čo eukaryoty sú mnohobunkové.
Porovnávacia charakteristika štruktúry buniek rastlín, živočíchov, baktérií a húb
Okrem znakov charakteristických pre prokaryoty a eukaryoty majú bunky rastlín, živočíchov, húb a baktérií aj množstvo znakov. Rastlinné bunky teda obsahujú špecifické organely - chloroplasty, ktoré určujú ich schopnosť fotosyntézy, zatiaľ čo tieto organely sa v iných organizmoch nenachádzajú. To samozrejme neznamená, že iné organizmy nie sú schopné fotosyntézy, pretože napríklad u baktérií sa vyskytuje na invagináciách plazmatickej membrány a jednotlivých membránových vezikúl v cytoplazme.
Rastlinné bunky spravidla obsahujú veľké vakuoly naplnené bunkovou šťavou. Nachádzajú sa aj v bunkách zvierat, húb a baktérií, ale majú úplne iný pôvod a plnia iné funkcie. Hlavnou rezervnou látkou nachádzajúcou sa vo forme pevných inklúzií v rastlinách je škrob, u zvierat a húb je to glykogén a v baktériách je to glykogén alebo volutín.
Ďalšou charakteristickou črtou týchto skupín organizmov je organizácia povrchového aparátu: bunky živočíšnych organizmov nemajú bunkovú stenu, ich plazmatická membrána je pokrytá len tenkou glykokalyxou, zatiaľ čo všetky ostatné ju majú. Je to úplne pochopiteľné, keďže spôsob kŕmenia zvierat je spojený so zachytávaním častíc potravy počas procesu fagocytózy a prítomnosť bunkovej steny by ich o túto príležitosť pripravila. Chemická povaha látky, ktorá tvorí bunkovú stenu, je v rôznych skupinách živých organizmov odlišná: ak je to v rastlinách celulóza, potom v hubách je to chitín a v baktériách je to mureín. Porovnávacia charakteristika štruktúry buniek rastlín, živočíchov, húb a baktérií
| Podpísať | Baktérie | Zvieratá | Huby | Rastliny |
| Spôsob výživy | Heterotrofné alebo autotrofné | Heterotrofný | Heterotrofný | Autotrofné |
| Organizácia dedičných informácií | Prokaryoty | Eukaryoty | Eukaryoty | Eukaryoty |
| lokalizácia DNA | Nukleoidy, plazmidy | Jadro, mitochondrie | Jadro, mitochondrie | Jadro, mitochondrie, plastidy |
| Plazmatická membrána | Jedzte | Jedzte | Jedzte | Jedzte |
| Bunková stena | Mureinovaya | — | Chitinous | Buničina |
| Cytoplazma | Jedzte | Jedzte | Jedzte | Jedzte |
| Organoidy | Ribozómy | Membránové a nemembránové, vrátane bunkového centra | Membránové a nemembránové | Membránové a nemembránové, vrátane plastidov |
| Organoidy pohybu | Bičíky a klky | Bičíky a mihalnice | Bičíky a mihalnice | Bičíky a mihalnice |
| Vacuoly | Málokedy | Kontraktilné, tráviace | Niekedy | Centrálna vakuola s bunkovou šťavou |
| Inklúzie | Glykogén, volutín | Glykogén | Glykogén | škrob |
Rozdiely v štruktúre buniek predstaviteľov rôznych kráľovstiev živej prírody sú znázornené na obrázku.

Chemické zloženie bunky. Makro- a mikroprvky. Vzťah medzi štruktúrou a funkciami anorganických a organických látok (bielkoviny, nukleové kyseliny, sacharidy, lipidy, ATP), ktoré tvoria bunku. Úloha chemikálií v bunke a ľudskom tele
Chemické zloženie bunky
Väčšina doteraz objavených chemických prvkov z periodickej tabuľky prvkov D. I. Mendelejeva bola nájdená v živých organizmoch. Jednak neobsahujú jediný prvok, ktorý by sa nenachádzal v neživej prírode a jednak sa ich koncentrácie v telách neživej prírody a živých organizmoch výrazne líšia.
Tieto chemické prvky tvoria anorganické a organické látky. Napriek skutočnosti, že v živých organizmoch prevládajú anorganické látky, sú to organické látky, ktoré určujú jedinečnosť ich chemického zloženia a fenomén života ako celku, pretože sú syntetizované hlavne organizmami v procese života a zohrávajú dôležitú úlohu v reakcie.

Veda študuje chemické zloženie organizmov a chemické reakcie, ktoré v nich prebiehajú. biochémia.
Je potrebné poznamenať, že obsah chemikálií v rôznych bunkách a tkanivách sa môže výrazne líšiť. Napríklad, ak v živočíšnych bunkách prevládajú bielkoviny medzi organickými zlúčeninami, potom v rastlinných bunkách prevládajú sacharidy.
| Chemický prvok | zemská kôra | Morská voda | Živé organizmy |
| O | 49.2 | 85.8 | 65-75 |
| C | 0.4 | 0.0035 | 15-18 |
| H | 1.0 | 10.67 | 8-10 |
| N | 0.04 | 0.37 | 1.5-3.0 |
| P | 0.1 | 0.003 | 0.20-1.0 |
| S | 0.15 | 0.09 | 0.15-0.2 |
| K | 2.35 | 0.04 | 0.15-0.4 |
| Ca | 3.25 | 0.05 | 0.04-2.0 |
| Cl | 0.2 | 0.06 | 0.05-0.1 |
| Mg | 2.35 | 0.14 | 0.02-0.03 |
| Na | 2.4 | 1.14 | 0.02-0.03 |
| Fe | 4.2 | 0.00015 | 0.01-0.015 |
| Zn | < 0.01 | 0.00015 | 0.0003 |
| Cu | < 0.01 | < 0.00001 | 0.0002 |
| ja | < 0.01 | 0.000015 | 0.0001 |
| F | 0.1 | 2.07 | 0.0001 |
Makro- a mikroprvky
V živých organizmoch sa nachádza asi 80 chemických prvkov, ale len 27 z týchto prvkov má svoje funkcie v bunke a organizme ustálené. Zvyšné prvky sú prítomné v malých množstvách a zjavne vstupujú do tela s jedlom, vodou a vzduchom. Obsah chemických prvkov v tele sa výrazne líši. Podľa koncentrácie sa delia na makroprvky a mikroprvky.
Koncentrácia každého z nich makronutrientov v tele presahuje 0,01% a ich celkový obsah je 99%. Medzi makroprvky patrí kyslík, uhlík, vodík, dusík, fosfor, síra, draslík, vápnik, sodík, chlór, horčík a železo. Prvé štyri z uvedených prvkov (kyslík, uhlík, vodík a dusík) sa tiež nazývajú organogénne, pretože sú súčasťou hlavných organických zlúčenín. Fosfor a síra sú tiež súčasťou mnohých organických látok, ako sú bielkoviny a nukleové kyseliny. Fosfor je nevyhnutný pre tvorbu kostí a zubov.
Bez zostávajúcich makroprvkov nie je možné normálne fungovanie tela. Draslík, sodík a chlór sa teda podieľajú na procesoch excitácie buniek. Draslík je tiež potrebný pre fungovanie mnohých enzýmov a zadržiavanie vody v bunke. Vápnik sa nachádza v bunkových stenách rastlín, kostiach, zuboch a schránkach mäkkýšov a je potrebný na kontrakciu svalových buniek a vnútrobunkový pohyb. Horčík je súčasťou chlorofylu, pigmentu, ktorý umožňuje priebeh fotosyntézy. Podieľa sa aj na biosyntéze bielkovín. Železo, okrem toho, že je súčasťou hemoglobínu, ktorý prenáša kyslík v krvi, je nevyhnutné pre procesy dýchania a fotosyntézy, ako aj pre fungovanie mnohých enzýmov.
Mikroelementy sú v organizme obsiahnuté v koncentráciách menších ako 0,01 % a ich celková koncentrácia v bunke nedosahuje 0,1 %. Medzi mikroelementy patrí zinok, meď, mangán, kobalt, jód, fluór atď. Zinok je súčasťou molekuly pankreatického hormónu inzulínu, meď je potrebná pre procesy fotosyntézy a dýchania. Kobalt je súčasťou vitamínu B12, ktorého nedostatok vedie k anémii. Jód je nevyhnutný pre syntézu hormónov štítnej žľazy, ktoré zabezpečujú normálny metabolizmus a fluorid je spojený s tvorbou zubnej skloviny.
Nedostatok aj nadbytok alebo narušenie metabolizmu makro- a mikroprvkov vedie k rozvoju rôznych chorôb. Najmä nedostatok vápnika a fosforu spôsobuje rachitu, nedostatok dusíka závažný nedostatok bielkovín, nedostatok železa anémiu a nedostatok jódu poruchu tvorby hormónov štítnej žľazy a zníženie rýchlosti metabolizmu. Zníženie príjmu fluoridov z vody a potravy do značnej miery podmieňuje narušenie obnovy zubnej skloviny a v dôsledku toho aj predispozíciu ku kazu. Olovo je toxické pre takmer všetky organizmy. Jeho nadbytok spôsobuje nezvratné poškodenie mozgu a centrálneho nervového systému, čo sa prejavuje stratou zraku a sluchu, nespavosťou, zlyhaním obličiek, kŕčmi a môže viesť aj k ochrnutiu a chorobám, ako je rakovina. Akútna otrava olovom je sprevádzaná náhlymi halucináciami a končí kómou a smrťou.
Nedostatok makro- a mikroprvkov možno kompenzovať zvýšením ich obsahu v potravinách a pitnej vode, ako aj užívaním liekov. Jód sa teda nachádza v morských plodoch a jódovanej soli, vápnik vo vaječných škrupinách atď.
Vzťah medzi štruktúrou a funkciami anorganických a organických látok (bielkoviny, nukleové kyseliny, sacharidy, lipidy, ATP), ktoré tvoria bunku. Úloha chemikálií v bunke a ľudskom tele
Anorganické látky
Chemické prvky bunky tvoria rôzne zlúčeniny – anorganické a organické. Anorganické látky bunky zahŕňajú vodu, minerálne soli, kyseliny atď., a organické látky zahŕňajú proteíny, nukleové kyseliny, sacharidy, lipidy, ATP, vitamíny atď.
Voda(H 2 O) je najbežnejšia anorganická látka bunky, ktorá má jedinečné fyzikálno-chemické vlastnosti. Nemá chuť, farbu ani vôňu. Hustota a viskozita všetkých látok sa hodnotí pomocou vody. Ako mnohé iné látky, aj voda môže existovať v troch skupenstvách agregácie: tuhá (ľad), kvapalná a plynná (para). Teplota topenia vody je $0°$С, bod varu je $100°$С, avšak rozpúšťanie iných látok vo vode môže tieto vlastnosti zmeniť. Pomerne vysoká je aj tepelná kapacita vody - 4200 kJ/mol K, čo jej dáva možnosť podieľať sa na termoregulačných procesoch. V molekule vody sú atómy vodíka umiestnené pod uhlom 105 ° $, zatiaľ čo zdieľané elektrónové páry sú odtiahnuté elektronegatívnejším atómom kyslíka. To určuje dipólové vlastnosti molekúl vody (jeden koniec je nabitý kladne a druhý záporne) a možnosť tvorby vodíkových väzieb medzi molekulami vody. Súdržnosť molekúl vody je základom javu povrchového napätia, vzlínavosti a vlastností vody ako univerzálneho rozpúšťadla. V dôsledku toho sa všetky látky delia na rozpustné vo vode (hydrofilné) a nerozpustné v nej (hydrofóbne). Vďaka týmto unikátnym vlastnostiam je predurčené, že voda sa stala základom života na Zemi.

Priemerný obsah vody v bunkách tela sa mení a môže sa meniť s vekom. V jeden a pol mesačnom ľudskom embryu teda obsah vody v bunkách dosahuje 97,5 %, u osemmesačného - 83 %, u novorodenca klesá na 74 % a v r. u dospelých je to v priemere 66 %. Telové bunky sa však líšia obsahom vody. Takže kosti obsahujú asi 20% vody, pečeň - 70% a mozog - 86%. Vo všeobecnosti sa to dá povedať koncentrácia vody v bunkách je priamo úmerná rýchlosti metabolizmu.
Minerálne soli môže byť v rozpustenom alebo nerozpustnom stave. Rozpustné soli disociovať na ióny – katióny a anióny. Najdôležitejšími katiónmi sú ióny draslíka a sodíka, ktoré uľahčujú prenos látok cez membránu a podieľajú sa na vzniku a vedení nervových vzruchov; ako aj vápenaté ióny, ktoré sa podieľajú na procesoch kontrakcie svalových vlákien a zrážania krvi; horčík, ktorý je súčasťou chlorofylu; železo, ktoré je súčasťou množstva bielkovín vrátane hemoglobínu. Najdôležitejšími aniónmi sú fosfátový anión, ktorý je súčasťou ATP a nukleových kyselín, a zvyšok kyseliny uhličitej, ktorý zjemňuje výkyvy pH prostredia. Ióny minerálnych solí zabezpečujú prienik vody samotnej do bunky a jej zadržiavanie v nej. Ak je koncentrácia soli v prostredí nižšia ako v bunke, potom voda preniká do bunky. Ióny určujú aj tlmiace vlastnosti cytoplazmy, teda jej schopnosť udržiavať konštantné mierne zásadité pH cytoplazmy, napriek neustálej tvorbe kyslých a zásaditých produktov v bunke.
Nerozpustné soli(CaCO 3, Ca 3 (PO 4) 2 atď.) sú súčasťou kostí, zubov, schránok a schránok jednobunkových a mnohobunkových živočíchov.
Okrem toho môžu organizmy produkovať ďalšie anorganické zlúčeniny, ako sú kyseliny a oxidy. Parietálne bunky ľudského žalúdka teda produkujú kyselinu chlorovodíkovú, ktorá aktivuje tráviaci enzým pepsín a oxid kremičitý preniká bunkovou stenou prasličky a vytvára schránky rozsievok. V posledných rokoch sa skúma aj úloha oxidu dusnatého (II) v signalizácii v bunkách a tele.
Organická hmota
Všeobecná charakteristika organických látok bunky
Organické látky bunky môžu byť reprezentované relatívne jednoduchými molekulami aj zložitejšími molekulami. V prípadoch, keď je komplexná molekula (makromolekula) tvorená značným počtom opakujúcich sa jednoduchších molekúl, ide o tzv. polymér a štrukturálne jednotky - monoméry. V závislosti od toho, či sa polymérne jednotky opakujú alebo nie, sú klasifikované ako pravidelné alebo nepravidelný. Polyméry tvoria až 90 % hmotnosti sušiny bunky. Patria do troch hlavných tried organických zlúčenín - sacharidy (polysacharidy), proteíny a nukleové kyseliny. Polysacharidy sú bežné polyméry, zatiaľ čo proteíny a nukleové kyseliny sú nepravidelné. V proteínoch a nukleových kyselinách je sekvencia monomérov mimoriadne dôležitá, pretože plnia informačnú funkciu.

Sacharidy
Sacharidy- Ide o organické zlúčeniny, ktoré sa skladajú najmä z troch chemických prvkov – uhlíka, vodíka a kyslíka, hoci množstvo uhľohydrátov obsahuje aj dusík alebo síru. Všeobecný vzorec uhľohydrátov je C m (H 2 O) n. Delia sa na jednoduché a zložité sacharidy.
Jednoduché sacharidy (monosacharidy) obsahujú jedinú molekulu cukru, ktorá sa nedá rozložiť na jednoduchšie. Ide o kryštalické látky, sladkej chuti a vysoko rozpustné vo vode. Monosacharidy sa aktívne podieľajú na bunkovom metabolizme a sú súčasťou komplexných sacharidov - oligosacharidov a polysacharidov.
Monosacharidy sú klasifikované podľa počtu atómov uhlíka (C3-C9), napr. pentózy(C 5) a hexózy(C6). Pentózy zahŕňajú ribózu a deoxyribózu. Ribóza je súčasťou RNA a ATP. deoxyribóza je súčasťou DNA. Hexózy (C 6 H 12 O 6) sú glukóza, fruktóza, galaktóza atď. Glukóza(hroznový cukor) sa nachádza vo všetkých organizmoch vrátane ľudskej krvi, pretože je energetickou rezervou. Je súčasťou mnohých komplexných cukrov: sacharózy, laktózy, maltózy, škrobu, celulózy atď. Fruktóza(ovocný cukor) sa nachádza v najvyšších koncentráciách v ovocí, mede a koreňoch cukrovej repy. Nielenže sa aktívne podieľa na metabolických procesoch, ale je tiež súčasťou sacharózy a niektorých polysacharidov, ako je inzulín.
Väčšina monosacharidov je schopná poskytnúť striebornú zrkadlovú reakciu a redukovať meď pri pridávaní tekutej kvapaliny (zmes roztokov síranu meďnatého a vínanu sodnodraselného) a varení.

TO oligosacharidy zahŕňajú sacharidy tvorené niekoľkými monosacharidovými zvyškami. Vo všeobecnosti sú tiež vysoko rozpustné vo vode a majú sladkú chuť. Podľa počtu týchto zvyškov sa rozlišujú disacharidy (dva zvyšky), trisacharidy (tri) atď.. Medzi disacharidy patrí sacharóza, laktóza, maltóza atď. Sacharóza(repný alebo trstinový cukor) pozostáva zo zvyškov glukózy a fruktózy, nachádza sa v zásobných orgánoch niektorých rastlín. Zvlášť veľa sacharózy je v okopaninách cukrovej repy a cukrovej trstiny, odkiaľ sa získavajú priemyselne. Slúži ako štandard pre sladkosť sacharidov. Laktóza, alebo mliečny cukor, tvorený zvyškami glukózy a galaktózy, sa nachádza v materskom a kravskom mlieku. maltóza(sladový cukor) pozostáva z dvoch glukózových jednotiek. Vzniká pri rozklade polysacharidov v semenách rastlín a v tráviacom systéme človeka a využíva sa pri výrobe piva.

Polysacharidy sú biopolyméry, ktorých monoméry sú mono- alebo disacharidové zvyšky. Väčšina polysacharidov je nerozpustná vo vode a má nesladenú chuť. Patria sem škrob, glykogén, celulóza a chitín. škrob- Ide o bielu práškovú hmotu, ktorá nie je zmáčaná vodou, ale po zaliatí horúcou vodou tvorí suspenziu - pastu. V skutočnosti sa škrob skladá z dvoch polymérov – menej rozvetvenej amylózy a viac rozvetveného amylopektínu (obr. 2.9). Monomérom amylózy aj amylopektínu je glukóza. Škrob je hlavnou zásobnou látkou rastlín, ktorá sa v obrovských množstvách hromadí v semenách, plodoch, hľuzách, pakorene a iných zásobných orgánoch rastlín. Kvalitatívna reakcia na škrob je reakcia s jódom, pri ktorej sa škrob sfarbí do modrofialova.

Glykogén(živočíšny škrob) je rezervný polysacharid živočíchov a húb, ktorý sa u ľudí hromadí v najväčšom množstve vo svaloch a pečeni. Je tiež nerozpustný vo vode a nemá sladkú chuť. Monomérom glykogénu je glukóza. V porovnaní s molekulami škrobu sú molekuly glykogénu ešte viac rozvetvené.
Celulóza, alebo celulóza, je hlavným podporným polysacharidom rastlín. Monomérom celulózy je glukóza. Nerozvetvené molekuly celulózy tvoria zväzky, ktoré tvoria súčasť bunkových stien rastlín. Celulóza je základom dreva, používa sa v stavebníctve, pri výrobe textílií, papiera, liehu a mnohých organických látok. Celulóza je chemicky inertná a nerozpúšťa sa v kyselinách ani zásadách. Nerozkladajú ho ani enzýmy v tráviacom systéme človeka, ale jeho trávenie uľahčujú baktérie v hrubom čreve. Okrem toho vláknina stimuluje kontrakcie stien gastrointestinálneho traktu, čím pomáha zlepšovať jeho fungovanie.

Chitin je polysacharid, ktorého monomér je monosacharid obsahujúci dusík. Je súčasťou bunkových stien húb a schránok článkonožcov. V ľudskom tráviacom systéme tiež chýba enzým na trávenie chitínu, majú ho len niektoré baktérie.
Funkcie uhľohydrátov. Sacharidy plnia v bunke plastové (stavebné), energetické, skladovacie a podporné funkcie. Tvoria bunkové steny rastlín a húb. Energetická hodnota štiepenia 1 g sacharidov je 17,2 kJ. Zásobnými látkami sú glukóza, fruktóza, sacharóza, škrob a glykogén. Sacharidy môžu byť tiež súčasťou komplexných lipidov a proteínov, tvoriacich glykolipidy a glykoproteíny, najmä v bunkových membránach. Nemenej dôležitá je úloha sacharidov pri medzibunkovom rozpoznávaní a vnímaní signálov z vonkajšieho prostredia, keďže fungujú ako receptory ako súčasť glykoproteínov.
Lipidy
Lipidy je chemicky heterogénna skupina látok s nízkou molekulovou hmotnosťou s hydrofóbnymi vlastnosťami. Tieto látky sú nerozpustné vo vode a tvoria v nej emulzie, ale sú vysoko rozpustné v organických rozpúšťadlách. Lipidy sú na dotyk mastné, mnohé z nich zanechávajú na papieri charakteristické nevysušujúce stopy. Spolu s bielkovinami a sacharidmi sú jednou z hlavných zložiek buniek. Obsah lipidov v rôznych bunkách nie je rovnaký, hlavne je ho veľa v semenách a plodoch niektorých rastlín, v pečeni, srdci, krvi.
V závislosti od štruktúry molekuly sa lipidy delia na jednoduché a zložité. TO jednoduché Lipidy zahŕňajú neutrálne lipidy (tuky), vosky a steroidy. Komplexné lipidy obsahujú aj ďalšiu, nelipidovú zložku. Najdôležitejšie z nich sú fosfolipidy, glykolipidy atď.
Tuky sú estery trojsýtneho alkoholu glycerolu a vyšších mastných kyselín. Väčšina mastných kyselín obsahuje 14-22 atómov uhlíka. Medzi nimi sú nasýtené aj nenasýtené, to znamená, že obsahujú dvojité väzby. Najbežnejšie nasýtené mastné kyseliny sú palmitová a stearová a najbežnejšie nenasýtené mastné kyseliny sú olejová. Niektoré nenasýtené mastné kyseliny sa v ľudskom tele nesyntetizujú alebo sú syntetizované v nedostatočnom množstve, a preto sú nevyhnutné. Glycerolové zvyšky tvoria hydrofilné „hlavy“ a zvyšky mastných kyselín tvoria hydrofóbne „chvosty“.

Tuky plnia predovšetkým zásobnú funkciu v bunkách a slúžia ako zdroj energie. Je na ne bohaté podkožné tukové tkanivo, ktoré plní tlmiace a tepelnoizolačné funkcie a u vodných živočíchov zvyšujú aj vztlak. Rastlinné tuky väčšinou obsahujú nenasýtené mastné kyseliny, v dôsledku čoho sú tekuté a tzv olejov. Oleje sú obsiahnuté v semenách mnohých rastlín, ako je slnečnica, sója, repka atď.
Vosky- Ide o estery a zmesi mastných kyselín a mastných alkoholov. U rastlín vytvárajú na povrchu listu film, ktorý chráni pred vyparovaním, prenikaním choroboplodných zárodkov a pod. U mnohých živočíchov pokrývajú telo alebo slúžia na stavbu plástov.
TO steroidy Patria sem lipidy, ako je cholesterol, základná zložka bunkových membrán, ako aj pohlavné hormóny estradiol, testosterón, vitamín D atď.
Fosfolipidy okrem zvyškov glycerolu a mastných kyselín obsahujú zvyšok kyseliny ortofosforečnej. Sú súčasťou bunkových membrán a zabezpečujú ich bariérové vlastnosti.
Glykolipidy sú tiež súčasťou membrán, ale ich obsah je tam malý. Nelipidovou časťou glykolipidov sú sacharidy.
Funkcie lipidov. Lipidy plnia v bunke plastickú (stavebnú), energetickú, zásobnú, ochrannú, vylučovaciu a regulačnú funkciu, okrem toho sú to vitamíny. Je nevyhnutnou súčasťou bunkových membrán. Pri odbúraní 1 g lipidov sa uvoľní 38,9 kJ energie. Sú uložené v rôznych orgánoch rastlín a živočíchov. Okrem toho podkožné tukové tkanivo chráni vnútorné orgány pred podchladením alebo prehriatím, ako aj pred šokom. Regulačná funkcia lipidov je spôsobená tým, že niektoré z nich sú hormóny. Tukové telo hmyzu slúži na vylučovanie.
Veveričky
Veveričky- Sú to vysokomolekulárne zlúčeniny, biopolyméry, ktorých monoméry sú aminokyseliny spojené peptidovými väzbami.
Aminokyselina nazývaná organická zlúčenina, ktorá má amino skupinu, karboxylovú skupinu a radikál. Celkovo sa v prírode nachádza asi 200 aminokyselín, ktoré sa líšia radikálmi a vzájomným usporiadaním funkčných skupín, no súčasťou bielkovín môže byť len 20 z nich. Tieto aminokyseliny sú tzv proteinogénne.

Bohužiaľ, nie všetky proteinogénne aminokyseliny sú v ľudskom tele syntetizované, preto sa delia na nahraditeľné a esenciálne. Neesenciálne aminokyseliny sa tvoria v ľudskom tele v požadovanom množstve, a nenahraditeľný- Nie. Musia byť dodávané potravou, ale môžu byť tiež čiastočne syntetizované črevnými mikroorganizmami. Úplne esenciálnych aminokyselín je 8. Patria sem valín, izoleucín, leucín, lyzín, metionín, treonín, tryptofán a fenylalanín. Napriek skutočnosti, že úplne všetky proteinogénne aminokyseliny sú syntetizované v rastlinách, rastlinné bielkoviny sú neúplné, pretože neobsahujú celú sadu aminokyselín a prítomnosť bielkovín vo vegetatívnych častiach rastlín zriedka presahuje 1-2% hmotnosti. . Preto je potrebné jesť bielkoviny nielen rastlinného, ale aj živočíšneho pôvodu.
Sekvencia dvoch aminokyselín spojených peptidovými väzbami sa nazýva dipeptid, z troch - tripeptid atď. Medzi peptidmi sú také dôležité zlúčeniny ako hormóny (oxytocín, vazopresín), antibiotiká atď. Reťazec viac ako dvadsiatich aminokyselín je tzv. polypeptid a polypeptidy obsahujúce viac ako 60 aminokyselinových zvyškov sú proteíny.

Úrovne štrukturálnej organizácie proteínov. Proteíny môžu mať primárne, sekundárne, terciárne a kvartérne štruktúry.
Primárna proteínová štruktúra- Toto lineárna sekvencia aminokyselín spojené peptidovou väzbou. Primárna štruktúra v konečnom dôsledku určuje špecifickosť proteínu a jeho jedinečnosť, pretože aj keď predpokladáme, že priemerný proteín obsahuje 500 aminokyselinových zvyškov, potom počet možných kombinácií je 20 500. Preto zmena v umiestnení aspoň jednej aminokyseliny kyseliny v primárnej štruktúre znamená zmenu sekundárnych a vyšších štruktúr, ako aj vlastností proteínu ako celku.

Štrukturálne vlastnosti proteínu určujú jeho priestorové usporiadanie - vznik sekundárnych a terciárnych štruktúr.
Sekundárna štruktúra predstavuje priestorové usporiadanie molekuly proteínu vo forme špirály alebo záhyby držané vodíkovými väzbami medzi atómami kyslíka a vodíka peptidových skupín rôznych závitov špirály alebo záhybov. Mnohé proteíny obsahujú viac či menej dlhé oblasti so sekundárnou štruktúrou. Sú to napríklad keratíny vlasov a nechtov, hodvábny fibroín.
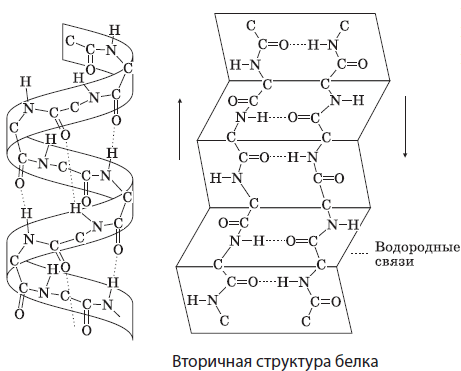
Terciárna štruktúra veverička ( globule) je tiež formou priestorového usporiadania polypeptidového reťazca držaného pohromade hydrofóbnymi, vodíkovými, disulfidovými (S-S) a inými väzbami. Je charakteristická pre väčšinu bielkovín v tele, ako je svalový myoglobín.

Kvartérna štruktúra- najzložitejšia, tvorená niekoľkými polypeptidovými reťazcami spojenými prevažne rovnakými väzbami ako v terciárnej (hydrofóbna, iónová a vodíková), ako aj inými slabými interakciami. Kvartérna štruktúra je charakteristická pre niekoľko proteínov, ako je hemoglobín, chlorofyl atď.

Na základe tvaru molekuly sa rozlišujú fibrilárne A guľovitý bielkoviny. Prvé z nich sú predĺžené, ako napríklad kolagén spojivového tkaniva alebo keratíny vlasov a nechtov. Globulárne bielkoviny majú tvar gule (globule), ako svalový myoglobín.
Jednoduché a zložité proteíny. Proteíny môžu byť jednoduché A komplexné. Jednoduché bielkoviny sa skladajú iba z aminokyselín, zatiaľ čo komplexné proteíny (lipoproteíny, chromoproteíny, glykoproteíny, nukleoproteíny atď.) obsahujú proteínové a neproteínové časti. Chromoproteíny obsahujú farebnú nebielkovinovú časť. Patria sem hemoglobín, myoglobín, chlorofyl, cytochrómy atď. V zložení hemoglobínu je teda každý zo štyroch polypeptidových reťazcov globínového proteínu spojený s neproteínovou časťou - hemom, v strede ktorého je železo ión, ktorý dáva hemoglobínu červenú farbu. Nebielkovinová časť lipoproteíny je lipid a glykoproteíny- uhľohydrát. Lipoproteíny aj glykoproteíny sú súčasťou bunkových membrán. Nukleoproteíny sú komplexy proteínov a nukleových kyselín (DNA a RNA). Vykonávajú najdôležitejšie funkcie v procesoch uchovávania a prenosu dedičných informácií.
Vlastnosti bielkovín. Mnohé bielkoviny sú vysoko rozpustné vo vode, existujú však aj také, ktoré sa rozpúšťajú len v roztokoch solí, zásad, kyselín alebo organických rozpúšťadiel. Štruktúra molekuly proteínu a jej funkčná aktivita závisí od podmienok prostredia. Strata jeho štruktúry molekulou proteínu pri zachovaní jeho primárnej štruktúry sa nazýva denaturácia.

K denaturácii dochádza v dôsledku zmien teploty, pH, atmosférického tlaku, vplyvom kyselín, zásad, solí ťažkých kovov, organických rozpúšťadiel a pod.. Obrátený proces obnovy sekundárnych a vyšších štruktúr je tzv. renaturácia nie je to však vždy možné. Úplné zničenie molekuly proteínu sa nazýva zničenie.
Funkcie proteínov. Proteíny plnia v bunke množstvo funkcií: plastickú (stavebnú), katalytickú (enzymatickú), energetickú, signalizačnú (receptorovú), kontraktilnú (motorickú), transportnú, ochrannú, regulačnú a zásobnú.
Konštrukčná funkcia proteínov je spojená s ich prítomnosťou v bunkových membránach a štruktúrnych zložkách bunky. Energia – vďaka tomu, že pri štiepení 1 g bielkovín sa uvoľní 17,2 kJ energie. Membránové receptorové proteíny sa aktívne podieľajú na vnímaní signálov z prostredia a ich prenose v bunke, ako aj na medzibunkovom rozpoznávaní. Bez bielkovín je pohyb buniek a organizmov ako celku nemožný, pretože tvoria základ bičíkov a mihalníc a tiež zabezpečujú svalovú kontrakciu a pohyb vnútrobunkových zložiek. V krvi ľudí a mnohých zvierat bielkovina hemoglobín prenáša kyslík a časť oxidu uhličitého, ostatné bielkoviny transportujú ióny a elektróny. Ochranná úloha proteínov je spojená predovšetkým s imunitou, pretože interferónový proteín je schopný ničiť mnohé vírusy a protilátkové proteíny potláčajú vývoj baktérií a iných cudzích látok. Medzi proteínmi a peptidmi je veľa hormónov, napríklad hormón pankreasu - inzulín, ktorý reguluje koncentráciu glukózy v krvi. V niektorých organizmoch môžu byť bielkoviny uložené ako rezervy, ako napríklad strukoviny v semenách alebo bielka kuracieho vajca.
Nukleové kyseliny
Nukleové kyseliny sú biopolyméry, ktorých monoméry sú nukleotidy. V súčasnosti sú známe dva typy nukleových kyselín: kyselina ribonukleová (RNA) a kyselina deoxyribonukleová (DNA).
Nukleotid tvorený dusíkatou zásadou, zvyškom pentózového cukru a zvyškom kyseliny ortofosforečnej. Charakteristiky nukleotidov sú určené hlavne dusíkatými bázami, ktoré ich tvoria, preto sa aj konvenčne označujú nukleotidy začiatočnými písmenami ich názvov. Nukleotidy môžu obsahovať päť dusíkatých báz: adenín (A), guanín (G), tymín (T), uracil (U) a cytozín (C). Pentózové nukleotidy - ribóza a deoxyribóza - určujú, ktorý nukleotid sa vytvorí - ribonukleotid alebo deoxyribonukleotid. Ribonukleotidy sú monoméry RNA, môžu pôsobiť ako signálne molekuly (cAMP) a sú súčasťou vysokoenergetických zlúčenín, ako je ATP, a koenzýmov, ako je NADP, NAD, FAD atď., a deoxyribonukleotidy sú súčasťou DNA.
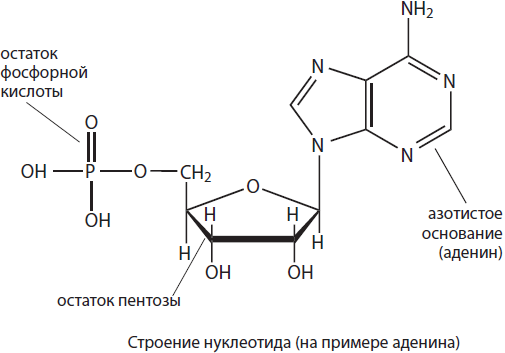
Kyselina deoxyribonukleová (DNA) je dvojvláknový biopolymér, ktorého monoméry sú deoxyribonukleotidy. Deoxyribonukleotidy obsahujú iba štyri dusíkaté bázy z piatich možných – adenín (A), tymín (T), guanín (G) alebo cytozín (C), ako aj zvyšky deoxyribózy a kyseliny ortofosforečnej. Nukleotidy v reťazci DNA sú navzájom spojené prostredníctvom zvyškov kyseliny ortofosforečnej, čím sa vytvorí fosfodiesterová väzba. Keď sa vytvorí dvojvláknová molekula, dusíkaté bázy sú nasmerované do vnútra molekuly. K spájaniu reťazcov DNA však nedochádza náhodne - dusíkaté bázy rôznych reťazcov sú navzájom spojené vodíkovými väzbami podľa princípu komplementarity: adenín je spojený s tymínom dvoma vodíkovými väzbami (A=T) a guanín je spojený s cytozínom tromi (G$≡C).

Boli jej nainštalované Chargaffove pravidlá:
- Počet nukleotidov DNA obsahujúcich adenín sa rovná počtu nukleotidov obsahujúcich tymín (A=T).
- Počet nukleotidov DNA obsahujúcich guanín sa rovná počtu nukleotidov obsahujúcich cytozín (G$≡$C).
- Súčet deoxyribonukleotidov obsahujúcich adenín a guanín sa rovná súčtu deoxyribonukleotidov obsahujúcich tymín a cytozín (A+G = T+C).
- Pomer súčtu deoxyribonukleotidov obsahujúcich adenín a tymín k súčtu deoxyribonukleotidov obsahujúcich guanín a cytozín závisí od typu organizmu.
Štruktúru DNA rozlúštili F. Crick a D. Watson (Nobelova cena za fyziológiu alebo medicínu, 1962). Podľa ich modelu je molekula DNA pravotočivá dvojzávitnica. Vzdialenosť medzi nukleotidmi v reťazci DNA je 0,34 nm.

Najdôležitejšou vlastnosťou DNA je schopnosť replikácie (samoduplikácie). Hlavnou funkciou DNA je uchovávanie a prenos dedičnej informácie, ktorá je zapísaná vo forme nukleotidových sekvencií. Stabilita molekuly DNA je udržiavaná výkonnými opravnými (obnovovacími) systémami, no ani tie nie sú schopné úplne eliminovať nepriaznivé vplyvy, čo v konečnom dôsledku vedie k vzniku mutácií. DNA eukaryotických buniek je sústredená v jadre, mitochondriách a plastidoch, zatiaľ čo v prokaryotických bunkách sa nachádza priamo v cytoplazme. Jadrová DNA je základom chromozómov, predstavujú ju otvorené molekuly. DNA mitochondrií, plastidov a prokaryotov je kruhová.
Ribonukleová kyselina (RNA)- biopolymér, ktorého monoméry sú ribonukleotidy. Obsahujú tiež štyri dusíkaté zásady - adenín (A), uracil (U), guanín (G) alebo cytozín (C), čím sa od DNA odlišujú jednou zo zásad (RNA namiesto tymínu obsahuje uracil). Pentózový cukorný zvyšok v ribonukleotidoch predstavuje ribóza. RNA sú väčšinou jednovláknové molekuly, s výnimkou niektorých vírusových. Existujú tri hlavné typy RNA: messenger alebo templát (mRNA), ribozomálna (rRNA) a transportná (tRNA). Všetky sa tvoria v procese prepisy- prepisovanie z molekúl DNA.
A RNA tvoria najmenšiu frakciu RNA v bunke (2-4%), čo je kompenzované ich diverzitou, keďže jedna bunka môže obsahovať tisíce rôznych mRNA. Sú to jednoreťazcové molekuly, ktoré sú templátmi na syntézu polypeptidových reťazcov. Informácie o štruktúre proteínu sú v nich zaznamenané vo forme nukleotidových sekvencií, pričom každá aminokyselina je kódovaná tripletom nukleotidov - kodón.
R RNA sú najrozšírenejším typom RNA v bunke (až 80 %). Ich molekulová hmotnosť je v priemere 3000-5000; vznikajú v jadierkach a sú súčasťou bunkových organel – ribozómov. Zdá sa, že rRNA tiež zohrávajú úlohu pri syntéze proteínov.
T RNA je najmenšia z molekúl RNA, pretože obsahuje iba 73-85 nukleotidov. Ich podiel na celkovom množstve RNA v bunke je asi 16 %. Funkciou tRNA je transport aminokyselín na miesto syntézy proteínov (ribozómy). Molekula tRNA má tvar ďatelinového listu. Na jednom konci molekuly je miesto pre pripojenie aminokyseliny a v jednej zo slučiek je triplet nukleotidov, komplementárnych ku kodónu mRNA a určujúcich, ktorú aminokyselinu bude tRNA niesť - antikodón.
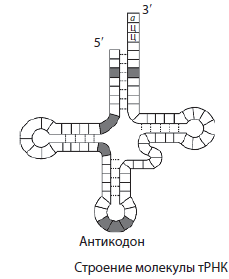
Všetky typy RNA sa aktívne podieľajú na procese implementácie dedičnej informácie, ktorá sa prepisuje z DNA na mRNA a tá vykonáva syntézu proteínov. tRNA dodáva aminokyseliny do ribozómov počas syntézy proteínov a rRNA je súčasťou samotných ribozómov.
Kyselina adenozíntrifosforečná (ATP) je nukleotid obsahujúci okrem dusíkatej bázy adenínu a ribózového zvyšku tri zvyšky kyseliny fosforečnej. Väzby medzi poslednými dvoma zvyškami fosforu sú vysokoenergetické (štiepením sa uvoľní 42 kJ/mol energie), pričom štandardnou chemickou väzbou pri štiepení vzniká 12 kJ/mol. Keď je potrebná energia, makroergická väzba ATP sa rozštiepi, vytvorí sa kyselina adenozíndifosforečná (ADP), zvyšok fosforu a uvoľní sa energia:
ATP + H20 $→$ ADP + H3PO4 + 42 kJ.
ADP sa môže tiež rozložiť za vzniku AMP (kyselina adenozínmonofosforečná) a zvyšku kyseliny fosforečnej:
ADP + H 2 O $→$ AMP + H 3 PO 4 + 42 kJ.
V procese energetického metabolizmu (pri dýchaní, fermentácii), ako aj v procese fotosyntézy, ADP viaže zvyšok fosforu a premieňa sa na ATP. Reakcia redukcie ATP je tzv fosforylácia. ATP je univerzálnym zdrojom energie pre všetky životné procesy živých organizmov.
Štúdium chemického zloženia buniek všetkých živých organizmov ukázalo, že obsahujú rovnaké chemické prvky, chemické látky, ktoré plnia rovnaké funkcie. Okrem toho v ňom bude fungovať časť DNA prenesená z jedného organizmu do druhého a proteín syntetizovaný baktériami alebo hubami bude v ľudskom tele vykonávať funkcie hormónu alebo enzýmu. Toto je jeden z dôkazov jednoty pôvodu organického sveta.

Bunková štruktúra. Vzťah medzi štruktúrou a funkciami častí a organel bunky je základom jej integrity
Bunková štruktúra
Štruktúra prokaryotických a eukaryotických buniek
Hlavnými štrukturálnymi zložkami buniek sú plazmatická membrána, cytoplazma a dedičný aparát. V závislosti od charakteristík organizácie sa rozlišujú dva hlavné typy buniek: prokaryotické a eukaryotické. Hlavným rozdielom medzi prokaryotickými bunkami a eukaryotickými bunkami je organizácia ich dedičného aparátu: u prokaryotov sa nachádza priamo v cytoplazme (táto oblasť cytoplazmy sa nazýva nukleoid) a nie je od nej oddelená membránovými štruktúrami, zatiaľ čo u eukaryotov je väčšina DNA sústredená v jadre, obklopenom dvojitou membránou. Okrem toho je genetická informácia prokaryotických buniek, umiestnených v nukleoide, zapísaná v kruhovej molekule DNA, zatiaľ čo v eukaryotoch sú molekuly DNA otvorené.
Na rozdiel od eukaryotov obsahuje cytoplazma prokaryotických buniek aj malý počet organel, zatiaľ čo eukaryotické bunky sa vyznačujú značnou rozmanitosťou týchto štruktúr.
Štruktúra a funkcie biologických membrán
Štruktúra biomembrány. Bunkové membrány a membránové organely eukaryotických buniek majú spoločné chemické zloženie a štruktúru. Zahŕňajú lipidy, bielkoviny a sacharidy. Membránové lipidy sú zastúpené najmä fosfolipidmi a cholesterolom. Väčšina membránových proteínov sú komplexné proteíny, ako sú glykoproteíny. Sacharidy sa v membráne nevyskytujú nezávisle, sú spojené s proteínmi a lipidmi. Hrúbka membrán je 7-10 nm.
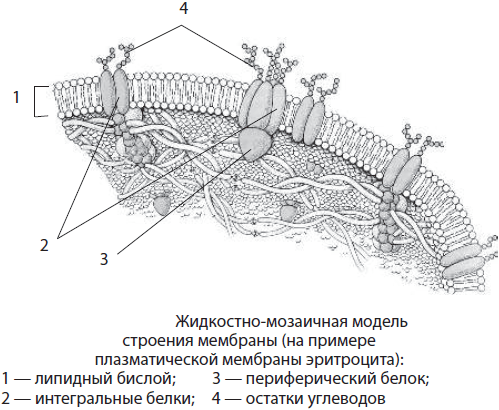
Podľa v súčasnosti všeobecne akceptovaného modelu fluidnej mozaiky štruktúry membrány tvoria lipidy dvojvrstvu, príp lipidová dvojvrstva, v ktorej hydrofilné „hlavy“ lipidových molekúl smerujú von a hydrofóbne „chvosty“ sú skryté vo vnútri membrány. Tieto „chvosty“ vďaka svojej hydrofóbnosti zabezpečujú oddelenie vodných fáz vnútorného prostredia bunky a jej prostredia. Proteíny sú spojené s lipidmi prostredníctvom rôznych typov interakcií. Niektoré proteíny sa nachádzajú na povrchu membrány. Takéto bielkoviny sa nazývajú periférne, alebo povrchný. Ostatné proteíny sú čiastočne alebo úplne ponorené do membrány – to sú integrálne, alebo ponorené proteíny. Membránové proteíny vykonávajú štrukturálne, transportné, katalytické, receptorové a iné funkcie.
Membrány nie sú ako kryštály, ich zložky sú neustále v pohybe, v dôsledku čoho sa medzi molekulami lipidov objavujú medzery - póry, cez ktoré môžu rôzne látky vstúpiť alebo opustiť bunku.
Biologické membrány sa líšia umiestnením v bunke, chemickým zložením a funkciami. Hlavné typy membrán sú plazmové a vnútorné. Plazmatická membrána obsahuje asi 45 % lipidov (vrátane glykolipidov), 50 % bielkovín a 5 % sacharidov. Nad povrch membrány vystupujú reťazce sacharidov, ktoré sú súčasťou komplexných proteínov-glykoproteínov a komplexných lipidov-glykolipidov. Plazmalemové glykoproteíny sú mimoriadne špecifické. Používajú sa napríklad na vzájomné rozpoznávanie buniek vrátane spermií a vajíčka.
Na povrchu živočíšnych buniek tvoria sacharidové reťazce tenkú povrchovú vrstvu - glykokalyx. Deteguje sa takmer vo všetkých živočíšnych bunkách, ale stupeň jeho expresie sa líši (10-50 µm). Glykokalyx poskytuje priamu komunikáciu medzi bunkou a vonkajším prostredím, kde dochádza k extracelulárnemu tráveniu; Receptory sú umiestnené v glykokalyxe. Okrem plazmalemy sú bunky baktérií, rastlín a húb tiež obklopené bunkovými membránami.
Vnútorné membrány eukaryotické bunky ohraničujú rôzne časti bunky a tvoria zvláštne „oddelenia“ - priehradky, ktorý podporuje oddelenie rôznych metabolických a energetických procesov. Môžu sa líšiť chemickým zložením a funkciami, ale ich všeobecný štruktúrny plán zostáva rovnaký.
Funkcie membrán:
- Obmedzujúce. Ide o to, že oddeľujú vnútorný priestor bunky od vonkajšieho prostredia. Membrána je polopriepustná, to znamená, že cez ňu môžu voľne prechádzať len tie látky, ktoré bunka potrebuje, a existujú mechanizmy na transport potrebných látok.
- Receptor. Primárne sa spája s vnímaním signálov prostredia a prenosom týchto informácií do bunky. Za túto funkciu sú zodpovedné špeciálne receptorové proteíny. Membránové proteíny sú tiež zodpovedné za bunkové rozpoznávanie podľa princípu „priateľ alebo nepriateľ“, ako aj za vytváranie medzibunkových spojení, z ktorých najviac skúmané sú synapsie nervových buniek.
- Katalytický. Na membránach sa nachádzajú početné komplexy enzýmov, v dôsledku čoho na nich prebiehajú intenzívne syntetické procesy.
- Transformácia energie. Súvisí s tvorbou energie, jej ukladaním vo forme ATP a spotrebou.
- Rozčlenenie. Membrány tiež vymedzujú priestor vo vnútri bunky, čím oddeľujú východiskové materiály reakcie a enzýmy, ktoré môžu vykonávať zodpovedajúce reakcie.
- Tvorba medzibunkových kontaktov. Napriek tomu, že hrúbka membrány je taká malá, že ju nemožno rozlíšiť voľným okom, slúži na jednej strane ako pomerne spoľahlivá bariéra pre ióny a molekuly, najmä vo vode rozpustné, a na druhej strane , zabezpečuje ich transport do bunky a von z bunky.
- Doprava.

Membránový transport. Vzhľadom na to, že bunky ako elementárne biologické systémy sú otvorené systémy, na zabezpečenie metabolizmu a energie, udržanie homeostázy, rastu, dráždivosti a ďalších procesov, je potrebný prenos látok cez membránu - membránový transport. V súčasnosti sa transport látok cez bunkovú membránu delí na aktívnu, pasívnu, endo- a exocytózu.
Pasívna doprava- Ide o typ transportu, ktorý prebieha bez spotreby energie z vyšších koncentrácií do nižších. V lipidoch rozpustné malé nepolárne molekuly (O 2, CO 2) ľahko prenikajú do bunky tým jednoduchá difúzia. Tie, ktoré nie sú rozpustné v lipidoch, vrátane malých nabitých častíc, sú zachytené nosnými proteínmi alebo prechádzajú špeciálnymi kanálmi (glukóza, aminokyseliny, K +, PO 4 3-). Tento druh pasívneho transportu je tzv uľahčená difúzia. Voda vstupuje do bunky cez póry v lipidovej fáze, ako aj cez špeciálne kanály vystlané proteínmi. Transport vody cez membránu je tzv osmózou.

Osmóza je v živote bunky mimoriadne dôležitá, pretože ak sa vloží do roztoku s vyššou koncentráciou solí ako v bunkovom roztoku, tak voda začne bunku opúšťať a objem živého obsahu sa začne zmenšovať. V živočíšnych bunkách sa bunka ako celok zmenšuje a v rastlinných bunkách cytoplazma zaostáva za bunkovou stenou, čo je tzv. plazmolýza. Keď je bunka umiestnená v roztoku menej koncentrovanom ako cytoplazma, dochádza k transportu vody v opačnom smere - do bunky. Rozšíriteľnosť cytoplazmatickej membrány má však limity a živočíšna bunka nakoniec praskne, zatiaľ čo rastlinná bunka to kvôli svojej silnej bunkovej stene neumožňuje. Fenomén naplnenia celého vnútorného priestoru bunky bunkovým obsahom sa nazýva tzv deplazmolýza. Pri príprave liekov, najmä na intravenózne podanie, je potrebné vziať do úvahy intracelulárnu koncentráciu solí, pretože to môže viesť k poškodeniu krvných buniek (na tento účel sa používa fyziologický roztok s koncentráciou 0,9% chloridu sodného). To je nemenej dôležité pri kultivácii buniek a tkanív, ako aj živočíšnych a rastlinných orgánov.

Aktívna doprava postupuje s výdajom energie ATP z nižšej koncentrácie látky na vyššiu. Vykonáva sa pomocou špeciálnych čerpacích proteínov. Proteíny pumpujú K +, Na +, Ca 2+ a ďalšie ióny cez membránu, čo podporuje transport základných organických látok, ako aj vznik nervových vzruchov atď.
Endocytóza- ide o aktívny proces absorpcie látok bunkou, pri ktorom membrána vytvára invaginácie a následne vytvára membránové vezikuly - fagozómy, ktoré obsahujú absorbované predmety. Potom sa primárny lyzozóm spojí s fagozómom a vytvorí sa sekundárny lyzozóm, alebo fagolyzozóm, alebo tráviaca vakuola. Obsah vezikuly je trávený lyzozómovými enzýmami a produkty rozpadu sú absorbované a asimilované bunkou. Nestrávené zvyšky sú z bunky odstránené exocytózou. Existujú dva hlavné typy endocytózy: fagocytóza a pinocytóza.
Fagocytóza je proces zachytávania bunkovým povrchom a absorpcie pevných častíc bunkou, a pinocytóza- tekutiny. Fagocytóza sa vyskytuje najmä v živočíšnych bunkách (jednobunkové živočíchy, ľudské leukocyty), zabezpečuje ich výživu a často chráni organizmus. Pinocytózou dochádza k absorpcii proteínov, komplexov antigén-protilátka počas imunitných reakcií atď. Mnohé vírusy však vstupujú do bunky aj pinocytózou alebo fagocytózou. V bunkách rastlín a húb je fagocytóza prakticky nemožná, pretože sú obklopené odolnými bunkovými membránami.

Exocytóza- proces obrátený k endocytóze. Z tráviacich vakuol sa tak uvoľňujú nestrávené zvyšky potravy a odstraňujú sa látky potrebné pre život bunky a organizmu ako celku. Napríklad k prenosu nervových impulzov dochádza v dôsledku uvoľnenia chemických poslov neurónom vysielajúcim impulz - mediátorov a v rastlinných bunkách sa takto vylučujú pomocné sacharidy bunkovej membrány.
Bunkové steny rastlinných buniek, húb a baktérií. Mimo membrány môže bunka vylučovať silnú štruktúru - bunková membrána, alebo bunková stena.
U rastlín je základom bunkovej steny celulóza, balené vo zväzkoch po 50-100 molekúl. Priestory medzi nimi sú vyplnené vodou a inými sacharidmi. Bunková stena rastliny je preniknutá tubulmi - plazmodesmata, cez ktorý prechádzajú membrány endoplazmatického retikula. Plazmodesmata vykonávajú transport látok medzi bunkami. K transportu látok, ako je voda, však môže dochádzať aj pozdĺž samotných bunkových stien. Postupom času sa v bunkovej stene rastlín hromadia rôzne látky vrátane tanínov alebo tukom podobných látok, čo vedie k lignifikácii alebo suberizácii samotnej bunkovej steny, vytláčaniu vody a odumieraniu bunkového obsahu. Medzi bunkovými stenami susedných rastlinných buniek sú rôsolovité rozpery - stredné platničky, ktoré ich držia pohromade a stmelujú telo rastliny ako celok. Zničia sa iba počas procesu dozrievania ovocia a pri opadaní listov.

Vytvárajú sa bunkové steny buniek húb chitín- uhľohydrát obsahujúci dusík. Sú dosť silné a sú vonkajšou kostrou bunky, no napriek tomu, podobne ako v rastlinách, zabraňujú fagocytóze.
V baktériách bunková stena obsahuje sacharidy s peptidovými fragmentmi - murein jeho obsah sa však medzi rôznymi skupinami baktérií výrazne líši. Iné polysacharidy sa môžu vylučovať aj na vrchnú časť bunkovej steny, čím sa vytvorí hlienová kapsula, ktorá chráni baktérie pred vonkajšími vplyvmi.
Membrána určuje tvar bunky, slúži ako mechanická podpora, plní ochrannú funkciu, zabezpečuje osmotické vlastnosti bunky, obmedzuje naťahovanie živého obsahu a zabraňuje prasknutiu bunky, ktoré sa zvyšuje v dôsledku vstupu vody . Navyše voda a látky v nej rozpustené prekonávajú bunkovú stenu pred vstupom do cytoplazmy alebo naopak pri jej výstupe, pričom voda sa cez bunkové steny transportuje rýchlejšie ako cez cytoplazmu.
Cytoplazma
Cytoplazma- Toto je vnútorný obsah bunky. Sú v nej ponorené všetky bunkové organely, jadro a rôzne odpadové produkty.
Cytoplazma spája všetky časti bunky navzájom a prebiehajú v nej početné metabolické reakcie. Cytoplazma je oddelená od prostredia a rozdelená na oddelenia membránami, to znamená, že bunky majú membránovú štruktúru. Môže byť v dvoch stavoch – sol a gél. Sol- ide o polotekutý rôsolovitý stav cytoplazmy, v ktorom prebiehajú životne dôležité procesy najintenzívnejšie a gél- hustejší, želatínový stav, ktorý bráni vzniku chemických reakcií a transportu látok.
Kvapalná časť cytoplazmy bez organel sa nazýva hyaloplazma. Hyaloplazma alebo cytosol je koloidný roztok, v ktorom je akási suspenzia pomerne veľkých častíc, napríklad proteínov, obklopených dipólmi molekúl vody. K zrážaniu tejto suspenzie nedochádza vďaka tomu, že majú rovnaký náboj a navzájom sa odpudzujú.
Organoidy
Organoidy- Sú to trvalé zložky bunky, ktoré vykonávajú špecifické funkcie.
V závislosti od štrukturálnych vlastností sa delia na membránové a nemembránové. Membrána organely sú zas klasifikované ako jednomembránové (endoplazmatické retikulum, Golgiho komplex a lyzozómy) alebo dvojmembránové (mitochondrie, plastidy a jadro). Bez membrány Organely sú ribozómy, mikrotubuly, mikrofilamenty a bunkové centrum. Z uvedených organel sú len ribozómy vlastné prokaryotom.
Štruktúra a funkcie jadra. Jadro- veľká dvojmembránová organela ležiaca v strede bunky alebo na jej okraji. Rozmery jadra sa môžu pohybovať od 3 do 35 mikrónov. Tvar jadra je najčastejšie guľovitý alebo elipsoidný, existujú však aj jadrá tyčinkovité, vretenovité, fazuľovité, laločnaté a dokonca aj segmentované. Niektorí vedci sa domnievajú, že tvar jadra zodpovedá tvaru samotnej bunky.
Väčšina buniek má jedno jadro, ale napríklad v bunkách pečene a srdca môžu byť dve a v množstve neurónov - až 15. Vlákna kostrového svalstva zvyčajne obsahujú veľa jadier, ale nie sú to bunky v plnom zmysle slova, keďže vznikajú ako výsledok splynutia viacerých buniek.
Jadro je obklopené jadrový obal a jeho vnútorný priestor je vyplnený jadrová šťava, alebo nukleoplazma (karyoplazma), do ktorej sú ponorené chromatín A jadierko. Jadro vykonáva také dôležité funkcie, ako je ukladanie a prenos dedičných informácií, ako aj riadenie života bunky.

Úloha jadra pri prenose dedičnej informácie bola presvedčivo preukázaná pri pokusoch so zelenou riasou Acetabularia. V jednej obrovskej bunke dosahujúcej dĺžku 5 cm sa rozlišuje čiapka, stopka a rizoid. Okrem toho obsahuje iba jedno jadro umiestnené v rizide. V 30. rokoch 20. storočia I. Hemmerling transplantoval jadro jedného druhu acetabularia so zelenou farbou do rizoidu iného druhu, s hnedou farbou, z ktorého bolo jadro odstránené. Rastlina s transplantovaným jadrom po určitom čase vyrástla nová čiapočka, podobne ako riasa darcu jadra. Zároveň čiapka alebo stopka, oddelená od rizoidu a neobsahujúca jadro, po určitom čase odumrela.

Jadrový obal tvorené dvoma membránami – vonkajšou a vnútornou, medzi ktorými je priestor. Medzimembránový priestor komunikuje s dutinou hrubého endoplazmatického retikula a vonkajšia membrána jadra môže niesť ribozómy. Jadrový obal je preniknutý početnými pórmi vystlanými špeciálnymi proteínmi. Transport látok prebieha cez póry: potrebné proteíny (vrátane enzýmov), ióny, nukleotidy a ďalšie látky vstupujú do jadra a molekuly RNA, vyčerpané proteíny a podjednotky ribozómov ho opúšťajú. Funkciou jadrového obalu je teda oddelenie obsahu jadra od cytoplazmy, ako aj regulácia metabolizmu medzi jadrom a cytoplazmou.
Nukleoplazma nazývaný obsah jadra, v ktorom je ponorený chromatín a jadierko. Je to koloidný roztok, chemicky pripomínajúci cytoplazmu. Enzýmy nukleoplazmy katalyzujú výmenu aminokyselín, nukleotidov, proteínov atď. Nukleoplazma je spojená s hyaloplazmou cez jadrové póry. Funkciou nukleoplazmy, podobne ako hyaloplazmy, je zabezpečiť prepojenie všetkých štruktúrnych zložiek jadra a uskutočniť množstvo enzymatických reakcií.
Chromatin nazývaný súbor tenkých filamentov a granúl ponorených do nukleoplazmy. Dá sa zistiť iba farbením, pretože indexy lomu chromatínu a nukleoplazmy sú približne rovnaké. Vláknitá zložka chromatínu je tzv euchromatínu a granulované - heterochromatín. Euchromatín je slabo zhutnený, pretože sa z neho číta dedičná informácia, zatiaľ čo viac špirálovitý heterochromatín je geneticky neaktívny.
Chromatín je štrukturálna modifikácia chromozómov v nedeliacom sa jadre. Chromozómy sú teda v jadre neustále prítomné, mení sa len ich stav v závislosti od funkcie, ktorú jadro práve plní.
Zloženie chromatínu zahŕňa predovšetkým nukleoproteínové proteíny (deoxyribonukleoproteíny a ribonukleoproteíny), ako aj enzýmy, z ktorých najdôležitejšie sú spojené so syntézou nukleových kyselín, a niektoré ďalšie látky.
Funkcie chromatínu spočívajú po prvé v syntéze nukleových kyselín špecifických pre daný organizmus, ktoré riadia syntézu špecifických proteínov, a po druhé, v prenose dedičných vlastností z materskej bunky na dcérske bunky, na čo slúži chromatínové vlákna sú počas procesu delenia zabalené do chromozómov.
Nucleolus- guľovité teleso, dobre viditeľné pod mikroskopom, s priemerom 1-3 mikróny. Vzniká na úsekoch chromatínu, v ktorých sú zakódované informácie o štruktúre rRNA a ribozomálnych proteínov. V jadre je často len jedno jadierko, ale v tých bunkách, kde prebiehajú intenzívne životne dôležité procesy, môžu byť jadierka dve alebo viac. Funkcie jadier sú syntéza rRNA a zostavenie ribozomálnych podjednotiek kombináciou rRNA s proteínmi pochádzajúcimi z cytoplazmy.
Mitochondrie- dvojmembránové organely okrúhleho, oválneho alebo tyčinkovitého tvaru, hoci sa nachádzajú aj špirálovité (v spermiách). Priemer mitochondrií je do 1 µm a dĺžka do 7 µm. Priestor vo vnútri mitochondrií je vyplnený matricou. Matrix- Toto je hlavná látka mitochondrií. V nej je ponorená kruhová molekula DNA a ribozómy. Vonkajšia membrána mitochondrií je hladká a nepriepustná pre mnohé látky. Vnútorná membrána má výstupky - cristas, čím sa zväčšuje povrch membrán pre chemické reakcie. Na povrchu membrány sú početné proteínové komplexy, ktoré tvoria takzvaný dýchací reťazec, ako aj enzýmy ATP syntetázy v tvare húb. Aeróbne štádium dýchania prebieha v mitochondriách, počas ktorého sa syntetizuje ATP.

Plastidy- veľké dvojmembránové organely, charakteristické len pre rastlinné bunky. Vnútorný priestor plastidov je vyplnený stroma, alebo matice. Stroma obsahuje viac či menej vyvinutý systém membránových vezikúl - tylakoidy, ktoré sa zhromažďujú v hromadách - zrná, ako aj vlastnú kruhovú molekulu DNA a ribozómy. Existujú štyri hlavné typy plastidov: chloroplasty, chromoplasty, leukoplasty a proplastidy.
Chloroplasty- sú to zelené plastidy s priemerom 3-10 mikrónov, jasne viditeľné pod mikroskopom. Nachádzajú sa len v zelených častiach rastlín – listoch, mladých stonkách, kvetoch a plodoch. Chloroplasty majú vo všeobecnosti oválny alebo elipsoidný tvar, ale môžu mať aj miskovitý, špirálovitý alebo dokonca laločnatý tvar. Počet chloroplastov v bunke je v priemere od 10 do 100 kusov. Avšak napríklad v niektorých riasach to môže byť jedna, mať výrazné rozmery a zložitý tvar – vtedy je tzv. chromatofór. V iných prípadoch môže počet chloroplastov dosiahnuť niekoľko stoviek, zatiaľ čo ich veľkosti sú malé. Farba chloroplastov je spôsobená hlavným pigmentom fotosyntézy - chlorofyl, aj keď obsahujú aj ďalšie pigmenty - karotenoidy. Karotenoidy sa prejavia až na jeseň, keď sa rozpadne chlorofyl v starnúcich listoch. Hlavnou funkciou chloroplastov je fotosyntéza. Svetelné reakcie fotosyntézy prebiehajú na tylakoidných membránach, na ktorých sú naviazané molekuly chlorofylu, a tmavé reakcie prebiehajú v stróme, kde sú obsiahnuté početné enzýmy.

Chromoplasty- Sú to žlté, oranžové a červené plastidy obsahujúce karotenoidné pigmenty. Tvar chromoplastov sa môže tiež výrazne líšiť: môžu byť rúrkové, guľovité, kryštalické atď. Chromoplasty dodávajú farbu kvetom a plodom rastlín, priťahujú opeľovače a rozdeľovače semien a plodov.
Leukoplasty- Sú to biele alebo bezfarebné plastidy, väčšinou okrúhleho alebo oválneho tvaru. Sú bežné v nefotosyntetických častiach rastlín, napríklad v šupke listov, hľúz zemiakov a pod.. Ukladajú živiny, najčastejšie škrob, ale v niektorých rastlinách to môžu byť bielkoviny alebo olej.
Plastidy sa tvoria v rastlinných bunkách z proplastidov, ktoré sú už prítomné v bunkách vzdelávacieho tkaniva a sú to malé dvojmembránové telieska. V počiatočných štádiách vývoja sa rôzne typy plastidov dokážu navzájom premieňať: keď sú vystavené svetlu, leukoplasty zemiakovej hľuzy a chromoplasty koreňa mrkvy zozelenajú.
Plastidy a mitochondrie sa nazývajú semiautonómne organely bunky, pretože majú svoje vlastné molekuly DNA a ribozómy, vykonávajú syntézu proteínov a delia sa nezávisle od delenia buniek. Tieto vlastnosti sú vysvetlené ich pôvodom z jednobunkových prokaryotických organizmov. „Nezávislosť“ mitochondrií a plastidov je však obmedzená, pretože ich DNA obsahuje príliš málo génov na voľnú existenciu, zatiaľ čo zvyšok informácií je zakódovaný v chromozómoch jadra, čo mu umožňuje kontrolovať tieto organely.
Endoplazmatické retikulum (ER), alebo endoplazmatické retikulum (ER), je jednomembránová organela, čo je sieť membránových dutín a tubulov zaberajúcich až 30 % obsahu cytoplazmy. Priemer EPS tubulov je asi 25-30 nm. Existujú dva typy EPS – drsný a hladký. Hrubý XPS nesie ribozómy, kde dochádza k syntéze bielkovín. Hladké XPS chýbajú mu ribozómy. Jeho funkciou je syntéza lipidov a sacharidov, ako aj transport, skladovanie a neutralizácia toxických látok. Vyvíja sa najmä v tých bunkách, kde prebiehajú intenzívne metabolické procesy, napríklad v pečeňových bunkách - hepatocytoch - a vláknach kostrového svalstva. Látky syntetizované v ER sú transportované do Golgiho aparátu. Zostavenie bunkových membrán sa vyskytuje aj v ER, ale ich tvorba je dokončená v Golgiho aparáte.
Golgiho aparát, alebo Golgiho komplex, je jednomembránová organela tvorená systémom plochých cisterien, od nich oddelených tubulov a vezikúl. Štrukturálna jednotka Golgiho aparátu je diktyozóm- hromada nádrží, z ktorých na jeden pól prichádzajú látky z EPS az opačného pólu sa po určitých premenách balia do vezikúl a posielajú do iných častí bunky. Priemer nádrží je asi 2 mikróny a priemer malých bublín je asi 20 až 30 mikrónov. Hlavnými funkciami Golgiho komplexu sú syntéza určitých látok a modifikácia (zmena) bielkovín, lipidov a sacharidov pochádzajúcich z ER, konečná tvorba membrán, ako aj transport látok v bunke, obnova jej štruktúr. a tvorbu lyzozómov. Golgiho aparát dostal svoje meno na počesť talianskeho vedca Camilla Golgiho, ktorý prvýkrát objavil túto organelu (1898).

lyzozómy- malé jednomembránové organely s priemerom do 1 μm, ktoré obsahujú hydrolytické enzýmy podieľajúce sa na vnútrobunkovom trávení. Membrány lyzozómov sú pre tieto enzýmy slabo priepustné, takže lyzozómy plnia svoje funkcie veľmi presne a cielene. Aktívne sa teda zúčastňujú procesu fagocytózy, tvoria tráviace vakuoly a v prípade hladovania alebo poškodenia niektorých častí bunky ich trávia bez toho, aby ovplyvnili ostatné. Nedávno bola objavená úloha lyzozómov v procesoch bunkovej smrti.
Vákuola je dutina v cytoplazme rastlinných a živočíšnych buniek, ohraničená membránou a vyplnená kvapalinou. V bunkách prvokov sa nachádzajú tráviace a kontraktilné vakuoly. Prvé sa podieľajú na procese fagocytózy, pretože rozkladajú živiny. Tie zabezpečujú udržiavanie rovnováhy voda-soľ vďaka osmoregulácii. U mnohobunkových živočíchov sa nachádzajú najmä tráviace vakuoly.
V rastlinných bunkách sú vakuoly vždy prítomné, sú obklopené špeciálnou membránou a naplnené bunkovou šťavou. Membrána obklopujúca vakuolu je svojím chemickým zložením, štruktúrou a funkciami podobná plazmatickej membráne. Bunková šťava je vodný roztok rôznych anorganických a organických látok, vrátane minerálnych solí, organických kyselín, uhľohydrátov, bielkovín, glykozidov, alkaloidov atď. Vakuola môže zaberať až 90% objemu bunky a vytláčať jadro na perifériu. Táto časť bunky plní funkciu zásobnú, vylučovaciu, osmotickú, ochrannú, lyzozomálnu a iné, keďže akumuluje živiny a odpadové látky, zabezpečuje prísun vody a udržiava tvar a objem bunky, obsahuje aj enzýmy na rozklad veľa bunkových komponentov. Okrem toho, biologicky aktívne látky vakuol môžu zabrániť mnohým zvieratám jesť tieto rastliny. V mnohých rastlinách dochádza v dôsledku napučiavania vakuol k rastu buniek predlžovaním.
Vakuoly sú prítomné aj v bunkách niektorých húb a baktérií, u húb však plnia len funkciu osmoregulácie, kým u siníc udržujú vztlak a podieľajú sa na procese asimilácie dusíka zo vzduchu.
Ribozómy- malé nemembránové organely s priemerom 15-20 mikrónov, pozostávajúce z dvoch podjednotiek - veľkej a malej. Eukaryotické ribozomálne podjednotky sú zostavené v jadierku a potom transportované do cytoplazmy. Ribozómy v prokaryotoch, mitochondriách a plastidoch sú menšie ako ribozómy v eukaryotoch. Ribozomálne podjednotky zahŕňajú rRNA a proteíny.

Počet ribozómov v bunke môže dosiahnuť niekoľko desiatok miliónov: v cytoplazme, mitochondriách a plastidoch sú vo voľnom stave a na hrubom ER - vo viazanom stave. Podieľajú sa na syntéze proteínov, najmä vykonávajú proces translácie - biosyntézu polypeptidového reťazca na molekule mRNA. Voľné ribozómy syntetizujú proteíny hyaloplazmy, mitochondrií, plastidov a svoje vlastné ribozomálne proteíny, zatiaľ čo ribozómy pripojené k hrubému ER vykonávajú transláciu proteínov na odstránenie z buniek, zostavenie membrán a tvorbu lyzozómov a vakuol.
Ribozómy sa môžu nachádzať jednotlivo v hyaloplazme alebo zostavené v skupinách počas simultánnej syntézy niekoľkých polypeptidových reťazcov na jednej mRNA. Takéto skupiny ribozómov sa nazývajú polyribozómy, alebo polyzómy.

Mikrotubuly- Sú to cylindrické duté nemembránové organely, ktoré prenikajú celou cytoplazmou bunky. Ich priemer je asi 25 nm, hrúbka steny je 6-8 nm. Sú tvorené množstvom proteínových molekúl tubulín, ktoré najskôr vytvoria 13 nití pripomínajúcich guľôčky a potom sa zostavia do mikrotubulu. Mikrotubuly tvoria cytoplazmatické retikulum, ktoré dáva bunke tvar a objem, spája plazmatickú membránu s ostatnými časťami bunky, zabezpečuje transport látok v bunke, podieľa sa na pohybe bunky a vnútrobunkových zložiek, ako aj na delenie genetického materiálu. Sú súčasťou bunkového centra a pohybových organel - bičíkov a mihalníc.
mikrovlákna, alebo mikrovlákna, sú tiež nemembránové organely, majú však vláknitý tvar a sú tvorené nie tubulínom, ale aktín. Zúčastňujú sa procesov membránového transportu, medzibunkového rozpoznávania, delenia bunkovej cytoplazmy a jej pohybu. Vo svalových bunkách interakcia aktínových mikrofilamentov s myozínovými vláknami sprostredkúva kontrakciu.
Mikrotubuly a mikrofilamenty tvoria vnútornú kostru bunky - cytoskelet. Ide o zložitú sieť vlákien, ktoré poskytujú mechanickú podporu plazmatickej membráne, určujú tvar bunky, umiestnenie bunkových organel a ich pohyb pri delení buniek.

Bunkové centrum- nemembránová organela nachádzajúca sa v živočíšnych bunkách v blízkosti jadra; v rastlinných bunkách chýba. Jeho dĺžka je približne 0,2-0,3 mikrónu a jeho priemer je 0,1-0,15 mikrónu. Bunkový stred tvoria dva centrioles, ležiace vo vzájomne kolmých rovinách, a žiarivá guľa z mikrotubulov. Každý centriol je tvorený deviatimi skupinami mikrotubulov, zhromaždených v skupinách po troch, t.j. tripletoch. Bunkové centrum sa zúčastňuje procesov zostavovania mikrotubulov, delenia bunkového dedičného materiálu, ako aj tvorby bičíkov a mihalníc.

Organely pohybu. Flagella A mihalnice Sú to bunkové výrastky pokryté plazmalemou. Základ týchto organel tvorí deväť párov mikrotubulov umiestnených pozdĺž periférie a dva voľné mikrotubuly v strede. Mikrotubuly sú vzájomne prepojené rôznymi proteínmi, zabezpečujúcimi ich koordinovanú odchýlku od osi - kmitanie. Oscilácie sú závislé od energie, to znamená, že na tento proces sa vynakladá energia vysokoenergetických väzieb ATP. Obnova stratených bičíkov a mihalníc je funkcia bazálnych teliesok, alebo kinetozómy nachádza na ich základni.
Dĺžka riasiniek je asi 10-15 nm a dĺžka bičíkov je 20-50 µm. Vplyvom prísne riadených pohybov bičíkov a mihalníc dochádza nielen k pohybu jednobunkových živočíchov, spermií a pod., ale aj k čisteniu dýchacích ciest a pohybu vajíčka vajíčkovodom, keďže všetky tieto časti ľudského tela sú lemované riasinkovým epitelom.

Inklúzie
Inklúzie- Sú to nestále zložky bunky, ktoré vznikajú a zanikajú počas jej života. Patria sem ako rezervné látky, napríklad zrnká škrobu alebo bielkovín v rastlinných bunkách, glykogénové granule v bunkách zvierat a húb, volutín v baktériách, kvapky tuku vo všetkých typoch buniek, tak aj odpadové produkty, najmä zvyšky potravín. nestrávené v dôsledku fagocytózy, tvoriace takzvané zvyškové telieska.

Vzťah medzi štruktúrou a funkciami častí a organel bunky je základom jej integrity
Každá z častí bunky je na jednej strane samostatnou štruktúrou so špecifickou štruktúrou a funkciami a na druhej strane je súčasťou komplexnejšieho systému nazývaného bunka. Väčšina dedičných informácií eukaryotickej bunky je sústredená v jadre, ale jadro samo o sebe nie je schopné zabezpečiť jej realizáciu, pretože na to je potrebná aspoň cytoplazma, ktorá pôsobí ako hlavná látka, a ribozómy, na ktorých k tejto syntéze dochádza. . Väčšina ribozómov sa nachádza na granulárnom endoplazmatickom retikule, odkiaľ sú proteíny najčastejšie transportované do Golgiho komplexu a následne po úprave do tých častí bunky, pre ktoré sú určené, prípadne sú vylučované. Membránové obaly proteínov a uhľohydrátov môžu byť vložené do membrán organel a cytoplazmatickej membrány, čím sa zabezpečí ich neustála obnova. Z Golgiho komplexu sa odpájajú aj lyzozómy a vakuoly, ktoré plnia dôležité funkcie. Napríklad bez lyzozómov by sa bunky rýchlo zmenili na akési smetisko pre odpadové molekuly a štruktúry.
Na uskutočnenie všetkých týchto procesov je potrebná energia produkovaná mitochondriami a v rastlinách chloroplastmi. A hoci sú tieto organely relatívne autonómne, keďže majú svoje vlastné molekuly DNA, niektoré z ich proteínov sú stále kódované jadrovým genómom a syntetizované v cytoplazme.
Bunka je teda neoddeliteľnou jednotou svojich základných zložiek, z ktorých každá plní svoju vlastnú jedinečnú funkciu.
Metabolizmus a premena energie sú vlastnosti živých organizmov. Energetický a plastový metabolizmus, ich vzťah. Etapy energetického metabolizmu. Fermentácia a dýchanie. Fotosyntéza, jej význam, kozmická úloha. Fázy fotosyntézy. Svetlé a tmavé reakcie fotosyntézy, ich vzťah. Chemosyntéza. Úloha chemosyntetických baktérií na Zemi
Metabolizmus a premena energie - vlastnosti živých organizmov
Bunku možno prirovnať k miniatúrnej chemickej továrni, v ktorej prebiehajú stovky a tisíce chemických reakcií.
Metabolizmus- súbor chemických premien zameraných na zachovanie a samorozmnožovanie biologických systémov.
Zahŕňa príjem látok do organizmu pri výžive a dýchaní, vnútrobunkový metabolizmus, príp metabolizmus, ako aj izoláciu konečných produktov metabolizmu.
Metabolizmus je neoddeliteľne spojený s procesmi premeny jedného druhu energie na iný. Napríklad v procese fotosyntézy sa svetelná energia ukladá vo forme energie chemických väzieb zložitých organických molekúl a počas procesu dýchania sa uvoľňuje a vynakladá sa na syntézu nových molekúl, mechanickú a osmotickú prácu, rozptýlené vo forme tepla atď.
Výskyt chemických reakcií v živých organizmoch je zabezpečený vďaka biologickým katalyzátorom proteínovej povahy - enzýmy, alebo enzýmy. Rovnako ako iné katalyzátory, aj enzýmy urýchľujú výskyt chemických reakcií v bunke desaťtisíckrát a niekedy ich dokonca umožňujú, ale nemenia povahu ani vlastnosti konečného produktu (produktov) reakcie a robia nezmenia sa. Enzýmy môžu byť jednoduché aj zložité bielkoviny, ktoré okrem bielkovinovej časti obsahujú aj nebielkovinovú časť – kofaktor (koenzým). Príkladmi enzýmov sú slinná amyláza, ktorá pri dlhšom žuvaní štiepi polysacharidy, a pepsín, ktorý zabezpečuje trávenie bielkovín v žalúdku.
Enzýmy sa líšia od neproteínových katalyzátorov vysokou špecifickosťou účinku, výrazným zvýšením reakčnej rýchlosti s ich pomocou, ako aj schopnosťou regulovať pôsobenie zmenou podmienok reakcie alebo interakciou rôznych látok s nimi. Okrem toho sa podmienky, za ktorých dochádza k enzymatickej katalýze, výrazne líšia od podmienok, za ktorých dochádza k neenzymatickej katalýze: optimálna teplota pre fungovanie enzýmov v ľudskom tele je 37 °C$, tlak by mal byť blízky atmosférickému tlaku a $pH$ prostredia môže výrazne zaváhať. Amyláza teda vyžaduje zásadité prostredie a pepsín vyžaduje kyslé prostredie.
Mechanizmus účinku enzýmov spočíva v znižovaní aktivačnej energie látok (substrátov), ktoré vstupujú do reakcie v dôsledku tvorby intermediárnych komplexov enzým-substrát.

Energetický a plastový metabolizmus, ich vzťah
Metabolizmus pozostáva z dvoch procesov prebiehajúcich súčasne v bunke: plastového a energetického metabolizmu.
Metabolizmus plastov (anabolizmus, asimilácia) je súbor syntéznych reakcií, ktoré zahŕňajú výdaj energie ATP. V procese metabolizmu plastov sa syntetizujú organické látky potrebné pre bunku. Príkladmi plastických výmenných reakcií sú fotosyntéza, biosyntéza proteínov a replikácia DNA (samoduplikácia).
Energetický metabolizmus (katabolizmus, disimilácia) je súbor reakcií, ktoré rozkladajú zložité látky na jednoduchšie. V dôsledku energetického metabolizmu sa energia uvoľňuje a ukladá vo forme ATP. Najdôležitejšie procesy energetického metabolizmu sú dýchanie a fermentácia.
Výmena plastov a energie sú neoddeliteľne spojené, pretože v procese výmeny plastov sa syntetizujú organické látky, čo si vyžaduje energiu ATP, a v procese výmeny energie sa organické látky rozkladajú a uvoľňuje sa energia, ktorá sa potom vynaloží na procesy syntézy. .
Organizmy prijímajú energiu v procese výživy, uvoľňujú ju a premieňajú do prístupnej formy najmä pri dýchaní. Podľa spôsobu výživy sa všetky organizmy delia na autotrofy a heterotrofy. Autotrofy schopné nezávisle syntetizovať organické látky z anorganických a heterotrofy používajte výhradne pripravené organické látky.
Etapy energetického metabolizmu
Napriek zložitosti reakcií energetického metabolizmu sa konvenčne delí na tri stupne: prípravný, anaeróbny (bez kyslíka) a aeróbny (kyslíkový).
Zapnuté prípravná fáza molekuly polysacharidov, lipidov, bielkovín, nukleových kyselín sa rozkladajú na jednoduchšie, napr. glukóza, glycerol a mastné kyseliny, aminokyseliny, nukleotidy atď. Toto štádium môže nastať priamo v bunkách alebo v črevách, odkiaľ sa odbúravajú páperové látky sú dodávané cez krvný obeh.
Anaeróbne štádium energetický metabolizmus je sprevádzaný ďalším rozkladom monomérov organických zlúčenín na ešte jednoduchšie medziprodukty, napríklad kyselinu pyrohroznovú alebo pyruvát. Nevyžaduje prítomnosť kyslíka a pre mnohé organizmy žijúce v bahne močiarov či v ľudských črevách je to jediný spôsob získavania energie. Anaeróbne štádium energetického metabolizmu prebieha v cytoplazme.
Rôzne látky môžu podliehať štiepeniu bez kyslíka, ale pomerne často je substrátom reakcií glukóza. Proces jeho bezkyslíkového štiepenia je tzv glykolýza. Pri glykolýze molekula glukózy stráca štyri atómy vodíka, t.j. oxiduje sa a vznikajú dve molekuly kyseliny pyrohroznovej, dve molekuly ATP a dve molekuly redukovaného nosiča vodíka $NADH + H^(+)$:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD -> 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
K tvorbe ATP z ADP dochádza v dôsledku priameho prenosu fosfátového aniónu z vopred fosforylovaného cukru a je tzv. fosforylácia substrátu.
Aeróbne štádium výmena energie môže prebiehať len v prítomnosti kyslíka, pričom medziprodukty vznikajúce pri bezkyslíkatom štiepení sa oxidujú na konečné produkty (oxid uhličitý a voda) a uvoľňuje sa väčšina energie uloženej v chemických väzbách organických zlúčenín. Premení sa na energiu vysokoenergetických väzieb 36 molekúl ATP. Táto etapa je tiež tzv tkanivové dýchanie. V neprítomnosti kyslíka sa medziproduktové zlúčeniny premieňajú na iné organické látky, procesom tzv fermentácia.
Dych
Mechanizmus bunkového dýchania je schematicky znázornený na obr.

Aeróbne dýchanie prebieha v mitochondriách, pričom kyselina pyrohroznová najskôr stráca jeden atóm uhlíka, čo je sprevádzané syntézou jedného redukčného ekvivalentu $NADH + H^(+)$ a molekuly acetylkoenzýmu A (acetyl-CoA):
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
Acetyl-CoA v mitochondriálnej matrici je zapojený do reťazca chemických reakcií, ktorých súhrn je tzv. Krebsov cyklus (cyklus trikarboxylovej kyseliny, cyklus kyseliny citrónovej). Počas týchto premien sa vytvoria dve molekuly ATP, acetyl-CoA sa úplne zoxiduje na oxid uhličitý a jeho vodíkové ióny a elektróny sa pridajú k nosičom vodíka $NADH + H^(+)$ a $FADH_2$. Nosiče transportujú vodíkové protóny a elektróny k vnútorným membránam mitochondrií, pričom vytvárajú cristae. Pomocou nosných proteínov sa vodíkové protóny pumpujú do medzimembránového priestoru a elektróny sa prenášajú cez takzvaný dýchací reťazec enzýmov umiestnených na vnútornej membráne mitochondrií a vypúšťajú sa na atómy kyslíka:
$O_2+2e^(-)→O_2^-$.
Treba poznamenať, že niektoré proteíny dýchacieho reťazca obsahujú železo a síru.
Z medzimembránového priestoru sú vodíkové protóny pomocou špeciálnych enzýmov - ATP syntáz transportované späť do mitochondriálnej matrice a uvoľnená energia sa v tomto prípade vynakladá na syntézu 34 molekúl ATP z každej molekuly glukózy. Tento proces sa nazýva Oxidačná fosforylácia. V mitochondriálnej matrici vodíkové protóny reagujú s kyslíkovými radikálmi za vzniku vody:
$4H^(+)+O_2^-→2H_2O$.
Súbor reakcií dýchania kyslíka možno vyjadriť takto:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
Celková rovnica dýchania vyzerá takto:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Fermentácia
Pri nedostatku kyslíka alebo jeho nedostatku nastáva fermentácia. Fermentácia je evolučne skorší spôsob získavania energie ako dýchanie, je však energeticky menej prospešný, pretože kvasením vznikajú organické látky, ktoré sú ešte energeticky bohaté. Existuje niekoľko hlavných typov fermentácie: kyselina mliečna, alkohol, kyselina octová atď. V kostrových svaloch sa teda v neprítomnosti kyslíka počas fermentácie kyselina pyrohroznová redukuje na kyselinu mliečnu, zatiaľ čo predtým vytvorené redukčné ekvivalenty sa spotrebúvajú a iba zostávajú dve molekuly ATP:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
Počas fermentácie pomocou kvasiniek sa kyselina pyrohroznová v prítomnosti kyslíka premieňa na etylalkohol a oxid uhoľnatý (IV):
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
Pri fermentácii pomocou mikroorganizmov môže z kyseliny pyrohroznovej vzniknúť aj kyselina octová, maslová, mravčia atď.
ATP, získaný ako výsledok energetického metabolizmu, sa v bunke vynakladá na rôzne druhy práce: chemickú, osmotickú, elektrickú, mechanickú a regulačnú. Chemická práca zahŕňa biosyntézu bielkovín, lipidov, sacharidov, nukleových kyselín a iných životne dôležitých zlúčenín. Osmotická práca zahŕňa procesy absorpcie bunkou a odstraňovania z nej látok, ktoré sú v extracelulárnom priestore v koncentráciách väčších ako v samotnej bunke. Elektrická práca úzko súvisí s osmotickou prácou, pretože pohybom nabitých častíc cez membrány sa vytvára membránový náboj a získavajú sa vlastnosti excitability a vodivosti. Mechanická práca zahŕňa pohyb látok a štruktúr vo vnútri bunky, ako aj bunky ako celku. Regulačná práca zahŕňa všetky procesy zamerané na koordináciu procesov v bunke.
Fotosyntéza, jej význam, kozmická úloha
Fotosyntéza je proces premeny svetelnej energie na energiu chemických väzieb organických zlúčenín za účasti chlorofylu.
V dôsledku fotosyntézy sa ročne vyprodukuje asi 150 miliárd ton organickej hmoty a približne 200 miliárd ton kyslíka. Tento proces zabezpečuje kolobeh uhlíka v biosfére, bráni hromadeniu oxidu uhličitého a tým zabraňuje skleníkovému efektu a prehrievaniu Zeme. Organické látky vznikajúce v dôsledku fotosyntézy nie sú úplne spotrebované inými organizmami, značná časť z nich v priebehu miliónov rokov vytvorila ložiská nerastných surovín (tvrdé a hnedé uhlie, ropa). V poslednom čase sa ako palivo začína používať aj repkový olej („bionafta“) a alkohol získaný z rastlinných zvyškov. Ozón sa tvorí z kyslíka pod vplyvom elektrických výbojov, ktoré tvoria ozónovú clonu, ktorá chráni všetok život na Zemi pred ničivými účinkami ultrafialových lúčov.
Náš krajan, vynikajúci rastlinný fyziológ K. A. Timiryazev (1843-1920), nazval úlohu fotosyntézy „kozmickou“, pretože spája Zem so Slnkom (vesmír) a zabezpečuje prílev energie na planétu.
Fázy fotosyntézy. Svetlé a tmavé reakcie fotosyntézy, ich vzťah
V roku 1905 anglický fyziológ rastlín F. Blackman zistil, že rýchlosť fotosyntézy sa nemôže zvyšovať donekonečna, nejaký faktor ju obmedzuje. Na základe toho predpokladal, že existujú dve fázy fotosyntézy: svetlo A tmavé. Pri nízkej intenzite svetla sa rýchlosť svetelných reakcií zvyšuje úmerne so zvyšovaním intenzity svetla a navyše tieto reakcie nezávisia od teploty, pretože nevyžadujú výskyt enzýmov. Svetelné reakcie sa vyskytujú na tylakoidných membránach.
Rýchlosť tmavých reakcií sa naopak zvyšuje so zvyšujúcou sa teplotou, avšak po dosiahnutí teplotného prahu $30°C$ sa tento nárast zastaví, čo naznačuje enzymatickú povahu týchto transformácií vyskytujúcich sa v stróme. Treba si uvedomiť, že na tmavé reakcie má určitý vplyv aj svetlo, napriek tomu, že sa im hovorí tmavé reakcie.

Svetelná fáza fotosyntézy sa vyskytuje na tylakoidných membránach nesúcich niekoľko typov proteínových komplexov, z ktorých hlavné sú fotosystémy I a II, ako aj ATP syntáza. Medzi fotosystémy patria pigmentové komplexy, ktoré okrem chlorofylu obsahujú aj karotenoidy. Karotenoidy zachytávajú svetlo v oblastiach spektra, kde chlorofyl nie, a tiež chránia chlorofyl pred zničením svetlom s vysokou intenzitou.
Súčasťou fotosystémov je okrem pigmentových komplexov aj množstvo akceptorových proteínov elektrónov, ktoré medzi sebou postupne prenášajú elektróny z molekúl chlorofylu. Sekvencia týchto proteínov je tzv elektrónový transportný reťazec chloroplastov.
S fotosystémom II je spojený aj špeciálny komplex bielkovín, ktorý zabezpečuje uvoľňovanie kyslíka pri fotosyntéze. Tento komplex uvoľňujúci kyslík obsahuje ióny mangánu a chlóru.
IN svetelná fáza svetelné kvantá alebo fotóny, dopadajúce na molekuly chlorofylu umiestnené na membránach tylakoidov, ich prenášajú do excitovaného stavu, vyznačujúceho sa vyššou energiou elektrónov. V tomto prípade sú excitované elektróny z chlorofylu fotosystému I prenesené cez reťazec medzičlánkov na vodíkový nosič NADP, ktorý pripája vodíkové protóny, vždy prítomné vo vodnom roztoku:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
Redukovaný $NADPH + H^(+)$ sa následne použije v tmavom štádiu. Elektróny z chlorofylu fotosystému II sa tiež prenášajú pozdĺž elektrónového transportného reťazca, ale zapĺňajú „elektrónové diery“ chlorofylu fotosystému I. Nedostatok elektrónov v chlorofyle fotosystému II je vyplnený odoberaním molekúl vody, ktoré prebieha za účasti komplexu uvoľňujúceho kyslík, ktorý už bol spomenutý vyššie. V dôsledku rozkladu molekúl vody, ktorý je tzv fotolýza, vznikajú vodíkové protóny a uvoľňuje sa molekulárny kyslík, ktorý je vedľajším produktom fotosyntézy:
$H_20 -> 2H^(+) + 2e^(-) + (1)/(2)0_2$.
Genetická informácia v bunke. Gény, genetický kód a jeho vlastnosti. Maticový charakter biosyntetických reakcií. Biosyntéza bielkovín a nukleových kyselín
Genetická informácia v bunke
Reprodukcia vlastného druhu je jednou zo základných vlastností živých vecí. Vďaka tomuto javu existuje podobnosť nielen medzi organizmami, ale aj medzi jednotlivými bunkami, ako aj ich organelami (mitochondrie a plastidy). Materiálnym základom tejto podobnosti je prenos genetickej informácie zašifrovanej v nukleotidovej sekvencii DNA, ktorý sa uskutočňuje prostredníctvom procesov replikácie DNA (samoduplikácie). Všetky vlastnosti a vlastnosti buniek a organizmov sa realizujú vďaka proteínom, ktorých štruktúra je primárne určená sekvenciou nukleotidov DNA. Preto má biosyntéza nukleových kyselín a proteínov prvoradý význam v metabolických procesoch. Štrukturálnou jednotkou dedičnej informácie je gén.
Gény, genetický kód a jeho vlastnosti
Dedičná informácia v bunke nie je monolitická, je rozdelená na samostatné „slová“ - gény.
Gene je elementárna jednotka genetickej informácie.
Práca na programe „Human Genome“, ktorý prebiehal súčasne vo viacerých krajinách a bol ukončený začiatkom tohto storočia, nám umožnila pochopiť, že človek má len asi 25-30 tisíc génov, ale informácie z väčšiny našej DNA sa nikdy nečíta, pretože obsahuje obrovské množstvo nezmyselných úsekov, opakovaní a génov kódujúcich črty, ktoré pre človeka stratili význam (chvost, ochlpenie na tele atď.). Okrem toho sa podarilo rozlúštiť množstvo génov zodpovedných za vývoj dedičných chorôb, ako aj cieľové gény liečiv. Praktická aplikácia výsledkov získaných pri realizácii tohto programu sa však odkladá, kým sa nerozlúštia genómy viacerých ľudí a neukáže sa, ako sa líšia.
Gény, ktoré kódujú primárnu štruktúru proteínovej, ribozomálnej alebo transferovej RNA, sa nazývajú štrukturálne a gény, ktoré zabezpečujú aktiváciu alebo potlačenie čítania informácií zo štrukturálnych génov - regulačné. Avšak aj štrukturálne gény obsahujú regulačné oblasti.
Dedičná informácia organizmov je zakódovaná v DNA vo forme určitých kombinácií nukleotidov a ich sekvencie - genetický kód. Jeho vlastnosti sú: tripletita, špecifickosť, univerzálnosť, redundancia a neprekrývanie sa. Okrem toho v genetickom kóde nie sú žiadne interpunkčné znamienka.
Každá aminokyselina je v DNA kódovaná tromi nukleotidmi - trojka, napríklad metionín je kódovaný tripletom TAC, to znamená, že kód je triplet. Na druhej strane každý triplet kóduje iba jednu aminokyselinu, čo je jeho špecifickosť alebo jednoznačnosť. Genetický kód je univerzálny pre všetky živé organizmy, to znamená, že dedičnú informáciu o ľudských bielkovinách dokážu prečítať baktérie a naopak. To naznačuje jednotu pôvodu organického sveta. Avšak 64 kombinácií troch nukleotidov zodpovedá iba 20 aminokyselinám, v dôsledku čoho môže byť jedna aminokyselina kódovaná 2-6 tripletmi, to znamená, že genetický kód je nadbytočný alebo degenerovaný. Tri triplety nemajú zodpovedajúce aminokyseliny, sú tzv stop kodóny, pretože označujú koniec syntézy polypeptidového reťazca.
Sekvencia báz v DNA tripletoch a aminokyseliny, ktoré kódujú

*Stop kodón označujúci koniec syntézy polypeptidového reťazca.
Skratky pre názvy aminokyselín:
Ala - alanín
Arg - arginín
Asn - asparagín
Asp - kyselina asparágová
Val - valín
Jeho - histidín
Gly - glycín
Gln - glutamín
Glu - kyselina glutámová
Ile - izoleucín
Leu - leucín
Liz - lyzín
pervitín - metionín
Pro - prolín
Ser - serín
Tyr - tyrozín
Tre - treonín
Tri - tryptofán
Fen - fenylalanín
Cis - cysteín
Ak začnete čítať genetickú informáciu nie z prvého nukleotidu v triplete, ale z druhého, tak sa nielen posunie čítací rámec, ale takto syntetizovaný proteín bude úplne odlišný nielen v sekvencii nukleotidov, ale aj v štruktúru a vlastnosti. Medzi trojčatami nie sú žiadne interpunkčné znamienka, takže neexistujú žiadne prekážky pri posúvaní čítacieho rámca, čím sa otvára priestor pre výskyt a udržiavanie mutácií.
Maticový charakter biosyntetických reakcií
Bakteriálne bunky sú schopné zdvojnásobiť sa každých 20-30 minút a eukaryotické bunky - každý deň a ešte častejšie, čo si vyžaduje vysokú rýchlosť a presnosť replikácie DNA. Okrem toho každá bunka obsahuje stovky a tisíce kópií mnohých proteínov, najmä enzýmov, preto je „kusový“ spôsob ich výroby pre ich reprodukciu neprijateľný. Progresívnejšou metódou je razenie, ktoré umožňuje získať početné presné kópie produktu a tiež znížiť jeho náklady. Na razenie je potrebná matrica, z ktorej sa vytvára odtlačok.
Princíp syntézy templátu v bunkách spočíva v tom, že nové molekuly proteínov a nukleových kyselín sa syntetizujú v súlade s programom zabudovaným do štruktúry už existujúcich molekúl rovnakých nukleových kyselín (DNA alebo RNA).
Biosyntéza bielkovín a nukleových kyselín
replikácia DNA. DNA je dvojvláknový biopolymér, ktorého monoméry sú nukleotidy. Ak by biosyntéza DNA prebiehala na princípe fotokopírovania, potom by nevyhnutne vznikli početné skreslenia a chyby v dedičnej informácii, ktoré by v konečnom dôsledku viedli k smrti nových organizmov. Preto proces zdvojenia DNA prebieha inak, polokonzervatívnym spôsobom: molekula DNA sa rozvinie a na každom z reťazcov sa syntetizuje nový reťazec podľa princípu komplementarity. Proces samoreprodukcie molekuly DNA, zabezpečujúci presné kopírovanie dedičnej informácie a jej prenos z generácie na generáciu, je tzv. replikácie(z lat. replikácie- opakovanie). V dôsledku replikácie sa vytvoria dve absolútne presné kópie materskej molekuly DNA, z ktorých každá nesie jednu kópiu materskej molekuly DNA.

Replikačný proces je v skutočnosti mimoriadne zložitý, keďže sa na ňom podieľa množstvo proteínov. Niektoré z nich rozvinú dvojitú špirálu DNA, iné prerušia vodíkové väzby medzi nukleotidmi komplementárnych reťazcov, iné (napríklad enzým DNA polymeráza) vyberú nové nukleotidy na princípe komplementarity atď. Dve molekuly DNA vytvorené ako výsledok replikácie sa počas delenia rozchádza na dve novovzniknuté dcérske bunky.
Chyby v procese replikácie sa vyskytujú veľmi zriedkavo, ale ak sa vyskytnú, sú veľmi rýchlo eliminované DNA polymerázami aj špeciálnymi opravnými enzýmami, pretože akákoľvek chyba v nukleotidovej sekvencii môže viesť k nezvratnej zmene štruktúry a funkcií proteínu. a v konečnom dôsledku nepriaznivo ovplyvňujú životaschopnosť novej bunky alebo dokonca jedinca.
Biosyntéza bielkovín. Ako obrazne povedal vynikajúci filozof 19. storočia F. Engels: „Život je formou existencie proteínových tiel.“ Štruktúra a vlastnosti proteínových molekúl sú určené ich primárnou štruktúrou, t.j. sekvenciou aminokyselín kódovaných v DNA. Od presnosti reprodukcie tejto informácie závisí nielen existencia samotného polypeptidu, ale aj fungovanie bunky ako celku, takže proces syntézy bielkovín má veľký význam. Zdá sa, že ide o najkomplexnejší proces syntézy v bunke, pretože zahŕňa až tristo rôznych enzýmov a iných makromolekúl. Navyše prúdi vysokou rýchlosťou, čo si vyžaduje ešte väčšiu presnosť.
V biosyntéze proteínov existujú dve hlavné fázy: transkripcia a translácia.
Prepis(z lat. prepis- prepisovanie) je biosyntéza molekúl mRNA na matrici DNA.

Keďže molekula DNA obsahuje dva antiparalelné reťazce, čítanie informácií z oboch reťazcov by viedlo k vytvoreniu úplne odlišných mRNA, preto je ich biosyntéza možná len na jednom z reťazcov, ktorý sa nazýva kódovací, alebo kodogénny, na rozdiel od druhého, nekódujúce alebo nekodogénne. Proces prepisovania zabezpečuje špeciálny enzým RNA polymeráza, ktorý vyberá RNA nukleotidy podľa princípu komplementarity. Tento proces sa môže vyskytnúť tak v jadre, ako aj v organelách, ktoré majú vlastnú DNA - mitochondrie a plastidy.
Molekuly mRNA syntetizované počas transkripcie prechádzajú zložitým procesom prípravy na transláciu (mitochondriálne a plastidové mRNA môžu zostať vo vnútri organel, kde nastáva druhá fáza biosyntézy bielkovín). Počas procesu dozrievania mRNA sa na ňu naviažu prvé tri nukleotidy (AUG) a chvost adenylových nukleotidov, ktorých dĺžka určuje, koľko kópií proteínu je možné syntetizovať na danej molekule. Až potom zrelé mRNA opúšťajú jadro cez jadrové póry.
Paralelne prebieha proces aktivácie aminokyselín v cytoplazme, počas ktorého sa aminokyselina spája so zodpovedajúcou voľnou tRNA. Tento proces je katalyzovaný špeciálnym enzýmom a vyžaduje ATP.
Vysielanie(z lat. vysielať- prenos) je biosyntéza polypeptidového reťazca na matrici mRNA, počas ktorej sa genetická informácia prekladá do sekvencie aminokyselín polypeptidového reťazca.

Druhá fáza syntézy proteínov sa najčastejšie vyskytuje v cytoplazme, napríklad na hrubom ER. Pre jeho výskyt je nevyhnutná prítomnosť ribozómov, aktivácia tRNA, počas ktorej pripájajú zodpovedajúce aminokyseliny, prítomnosť Mg2+ iónov, ako aj optimálne podmienky prostredia (teplota, pH, tlak a pod.).
Ak chcete spustiť vysielanie ( zasvätenie) malá ribozomálna podjednotka sa pripojí k molekule mRNA pripravenej na syntézu a potom sa podľa princípu komplementarity k prvému kodónu (AUG) vyberie tRNA nesúca aminokyselinu metionín. Až potom sa pripojí veľká ribozomálna podjednotka. V rámci zostaveného ribozómu sú dva kodóny mRNA, z ktorých prvý je už obsadený. Druhá tRNA, ktorá tiež nesie aminokyselinu, sa pridá ku kodónu, ktorý s ňou susedí, a potom sa pomocou enzýmov vytvorí peptidová väzba medzi aminokyselinovými zvyškami. Ribozóm presúva jeden kodón mRNA; prvá tRNA uvoľnená z aminokyseliny sa vracia do cytoplazmy po ďalšej aminokyseline a fragment budúceho polypeptidového reťazca visí akoby na zostávajúcej tRNA. Ďalšia tRNA sa pripojí k novému kodónu, ktorý sa ocitne v ribozóme, proces sa opakuje a krok za krokom sa polypeptidový reťazec predlžuje, t.j. predĺženie.
Koniec syntézy bielkovín ( ukončenie) sa vyskytuje hneď, ako sa v molekule mRNA stretne špecifická nukleotidová sekvencia, ktorá nekóduje aminokyselinu (stop kodón). Potom sa ribozóm, mRNA a polypeptidový reťazec oddelia a novosyntetizovaný proteín získa príslušnú štruktúru a je transportovaný do časti bunky, kde bude vykonávať svoje funkcie.
Translácia je energeticky veľmi náročný proces, pretože energia jednej molekuly ATP sa spotrebuje na pripojenie jednej aminokyseliny k tRNA a niekoľko ďalších sa používa na pohyb ribozómu pozdĺž molekuly mRNA.
Na urýchlenie syntézy určitých proteínových molekúl sa môže na molekulu mRNA postupne pripojiť niekoľko ribozómov, ktoré tvoria jednu štruktúru - polyzóm.
Bunka je genetická jednotka živej veci. Chromozómy, ich štruktúra (tvar a veľkosť) a funkcie. Počet chromozómov a ich druhová stálosť. Somatické a zárodočné bunky. Životný cyklus bunky: interfáza a mitóza. Mitóza je delenie somatických buniek. meióza. Fázy mitózy a meiózy. Vývoj zárodočných buniek u rastlín a živočíchov. Bunkové delenie je základom pre rast, vývoj a rozmnožovanie organizmov. Úloha meiózy a mitózy
Bunka je genetická jednotka živej veci.
Napriek tomu, že nukleové kyseliny sú nositeľmi genetickej informácie, implementácia tejto informácie je mimo bunky nemožná, čo sa dá ľahko dokázať na príklade vírusov. Tieto organizmy, ktoré často obsahujú len DNA alebo RNA, sa nedokážu samostatne rozmnožovať, na to musia využiť dedičný aparát bunky. Nedokážu ani preniknúť do bunky bez pomoci samotnej bunky, s výnimkou použitia membránových transportných mechanizmov alebo v dôsledku poškodenia bunky. Väčšina vírusov je nestabilná, umierajú už po niekoľkých hodinách vystavenia na čerstvom vzduchu. Bunka je teda genetická jednotka živej bytosti, ktorá má minimálny súbor komponentov na uchovávanie, zmenu a implementáciu dedičnej informácie, ako aj jej prenos na potomkov.
Väčšina genetickej informácie eukaryotickej bunky sa nachádza v jadre. Zvláštnosťou jej organizácie je, že na rozdiel od DNA prokaryotickej bunky nie sú molekuly DNA eukaryot uzavreté a tvoria zložité komplexy s proteínmi – chromozómami.
Chromozómy, ich štruktúra (tvar a veľkosť) a funkcie
Chromozóm(z gréčtiny chróm- farba, sfarbenie a soma- telo) je štruktúra bunkového jadra, ktorá obsahuje gény a nesie určité dedičné informácie o vlastnostiach a vlastnostiach organizmu.
Niekedy sa kruhové molekuly DNA prokaryotov nazývajú aj chromozómy. Chromozómy sú schopné autoduplikácie, majú štrukturálnu a funkčnú individualitu a zachovávajú si ju po celé generácie. Každá bunka nesie všetky dedičné informácie tela, no funguje v nej len malá časť.
Základom chromozómu je dvojvláknová molekula DNA nabitá proteínmi. U eukaryotov interagujú histónové a nehistónové proteíny s DNA, zatiaľ čo u prokaryotov histónové proteíny chýbajú.
Chromozómy sú najlepšie viditeľné pod svetelným mikroskopom počas delenia buniek, keď v dôsledku zhutnenia nadobúdajú vzhľad tyčovitých teliesok oddelených primárnym zúžením - centroméra — na ramenách. Na chromozóme môže byť tiež sekundárne zúženie, ktorý v niektorých prípadoch oddeľuje tzv satelit. Konce chromozómov sa nazývajú teloméry. Teloméry zabraňujú zlepeniu koncov chromozómov a zabezpečujú ich pripojenie k jadrovej membráne v nedeliacej sa bunke. Na začiatku delenia sú chromozómy zdvojené a pozostávajú z dvoch dcérskych chromozómov - chromatid, upevnený na centromére.

Podľa tvaru sa chromozómy delia na rovnoramenné, nerovnoramenné a tyčinkovité chromozómy. Veľkosti chromozómov sa výrazne líšia, ale priemerný chromozóm má rozmery 5 $ × $ 1,4 mikrónov.
V niektorých prípadoch chromozómy v dôsledku početných duplikácií DNA obsahujú stovky a tisíce chromatidov: takéto obrovské chromozómy sa nazývajú polyetylén. Nachádzajú sa v slinných žľazách lariev Drosophila, ako aj v tráviacich žľazách škrkaviek.
Počet chromozómov a ich druhová stálosť. Somatické a zárodočné bunky
Podľa bunkovej teórie je bunka jednotkou štruktúry, životnej činnosti a vývoja organizmu. Na bunkovej úrovni sú teda zabezpečené také dôležité funkcie živých vecí, ako je rast, reprodukcia a vývoj organizmu. Bunky mnohobunkových organizmov možno rozdeliť na somatické a reprodukčné bunky.
Somatické bunky- sú to všetky bunky tela vytvorené ako výsledok mitotického delenia.
Štúdium chromozómov umožnilo zistiť, že somatické bunky tela každého biologického druhu sa vyznačujú konštantným počtom chromozómov. Napríklad človek ich má 46. Súbor chromozómov somatických buniek je tzv diploidný(2n), alebo dvojité.

Pohlavné bunky, alebo gaméty, sú špecializované bunky používané na sexuálne rozmnožovanie.
Gaméty obsahujú vždy o polovicu menej chromozómov ako somatické bunky (u ľudí - 23), preto sa súbor chromozómov zárodočných buniek nazýva tzv. haploidný(n), alebo slobodný. Jeho tvorba je spojená s delením meiotických buniek.
Množstvo DNA v somatických bunkách je označené ako 2c a v pohlavných bunkách - 1c. Genetický vzorec somatických buniek je napísaný ako 2n2c a sexuálnych buniek - 1n1c.
V jadrách niektorých somatických buniek sa počet chromozómov môže líšiť od ich počtu v somatických bunkách. Ak je tento rozdiel väčší ako jedna, dve, tri atď. haploidné množiny, potom sa takéto bunky nazývajú polyploidný(tri-, tetra-, pentaploid, v tomto poradí). V takýchto bunkách zvyčajne prebiehajú metabolické procesy veľmi intenzívne.
Počet chromozómov sám o sebe nie je druhovo špecifickým znakom, pretože rôzne organizmy môžu mať rovnaký počet chromozómov, ale príbuzné môžu mať rôzny počet. Napríklad malarické plazmodium a škrkavka konská majú každý dva chromozómy, zatiaľ čo ľudia a šimpanzy majú 46 a 48.
Ľudské chromozómy sú rozdelené do dvoch skupín: autozómy a pohlavné chromozómy (heterochromozómy). Autosome v ľudských somatických bunkách je 22 párov, sú rovnaké pre mužov a ženy, a pohlavné chromozómy len jeden pár, ale práve ten určuje pohlavie jedinca. Existujú dva typy pohlavných chromozómov - X a Y. Bunky ženského tela nesú dva chromozómy X a mužské - X a Y.
karyotyp- ide o súbor vlastností chromozómovej sady organizmu (počet chromozómov, ich tvar a veľkosť).
Podmienený záznam karyotypu zahŕňa celkový počet chromozómov, pohlavných chromozómov a možné odchýlky v sade chromozómov. Napríklad karyotyp normálneho muža sa zapíše ako 46, XY a karyotyp normálnej ženy je 46, XX.
Životný cyklus bunky: interfáza a mitóza
Bunky nevznikajú zakaždým nanovo, vznikajú až v dôsledku delenia materských buniek. Dcérske bunky po rozdelení potrebujú určitý čas na vytvorenie organel a získanie vhodnej štruktúry, ktorá by zabezpečila výkon špecifickej funkcie. Toto časové obdobie sa nazýva zrenia.
Nazýva sa časový úsek od objavenia sa bunky v dôsledku delenia až po jej rozdelenie alebo smrť životný cyklus bunky.
V eukaryotických bunkách je životný cyklus rozdelený na dve hlavné fázy: interfázu a mitózu.

Medzifáza- ide o časový úsek životného cyklu, počas ktorého sa bunka nedelí a funguje normálne. Interfáza je rozdelená do troch periód: G 1 -, S- a G 2 - periódy.
G1 -obdobie(presyntetické, postmitotické) je obdobie bunkového rastu a vývoja, počas ktorého dochádza k aktívnej syntéze RNA, proteínov a ďalších látok potrebných na úplnú podporu života novovytvorenej bunky. Ku koncu tohto obdobia sa bunka môže začať pripravovať na duplikáciu svojej DNA.
IN S-obdobie(syntetické) dochádza k samotnému procesu replikácie DNA. Jedinou časťou chromozómu, ktorá neprechádza replikáciou, je centroméra, takže výsledné molekuly DNA sa úplne nerozchádzajú, ale zostávajú v nej držané pohromade a na začiatku delenia má chromozóm vzhľad v tvare X. Genetický vzorec bunky po zdvojnásobení DNA je 2n4c. Aj v S-období sú centrioly bunkového centra zdvojené.
G2 -obdobie(postsyntetický, premitotický) sa vyznačuje intenzívnou syntézou RNA, proteínov a ATP nevyhnutných pre proces bunkového delenia, ako aj separáciu centriol, mitochondrií a plastidov. Až do konca interfázy zostáva chromatín a jadierko jasne rozlíšiteľné, integrita jadrového obalu nie je narušená a organely sa nemenia.
Niektoré z telesných buniek sú schopné vykonávať svoje funkcie počas celého života tela (neuróny nášho mozgu, svalové bunky srdca), iné existujú len krátky čas, po ktorom odumierajú (bunky črevného epitelu, epidermálne bunky koža). V dôsledku toho musí telo neustále podstupovať procesy delenia buniek a tvorby nových, ktoré by nahradili odumreté. Bunky schopné deliť sa tzv stonka. V ľudskom tele sa nachádzajú v červenej kostnej dreni, v hlbokých vrstvách epidermis kože a na iných miestach. Pomocou týchto buniek môžete pestovať nový orgán, dosiahnuť omladenie a tiež klonovať telo. Perspektívy použitia kmeňových buniek sú úplne jasné, ale o morálnych a etických aspektoch tohto problému sa stále diskutuje, pretože vo väčšine prípadov sa používajú embryonálne kmeňové bunky získané z ľudských embryí usmrtených pri potrate.
Trvanie interfázy v rastlinných a živočíšnych bunkách je v priemere 10-20 hodín, zatiaľ čo mitóza trvá asi 1-2 hodiny.
Počas postupných delení v mnohobunkových organizmoch sa dcérske bunky stávajú čoraz rozmanitejšími, pretože čítajú informácie z rastúceho počtu génov.
Niektoré bunky sa časom prestanú deliť a odumierajú, čo môže byť spôsobené dokončením určitých funkcií, ako v prípade epidermálnych kožných buniek a krviniek, alebo poškodením týchto buniek environmentálnymi faktormi, najmä patogénmi. Geneticky naprogramovaná bunková smrť je tzv apoptóza, zatiaľ čo náhodná smrť - nekróza.
Mitóza je delenie somatických buniek. Fázy mitózy
Mitóza- metóda nepriameho delenia somatických buniek.
Počas mitózy bunka prechádza sériou po sebe nasledujúcich fáz, v dôsledku ktorých každá dcérska bunka dostane rovnakú sadu chromozómov ako materská bunka.
Mitóza je rozdelená do štyroch hlavných fáz: profáza, metafáza, anafáza a telofáza. Profáza- najdlhšia fáza mitózy, počas ktorej kondenzuje chromatín, výsledkom čoho sú chromozómy v tvare X pozostávajúce z dvoch chromatíd (dcérskych chromozómov), ktoré sa stávajú viditeľnými. V tomto prípade nukleolus zmizne, centrioly sa rozchádzajú k pólom bunky a z mikrotubulov sa začne vytvárať achromatínové vreteno (deliace vreteno). Na konci profázy sa jadrová membrána rozpadne na samostatné vezikuly.

IN metafáza Chromozómy sú usporiadané pozdĺž rovníka bunky so svojimi centromérmi, ku ktorým sú pripojené mikrotubuly plne vytvoreného vretienka. V tomto štádiu delenia sú chromozómy najviac zhutnené a majú charakteristický tvar, ktorý umožňuje študovať karyotyp.
IN anafázy Rýchla replikácia DNA nastáva v centroméroch, v dôsledku čoho sú chromozómy rozdelené a chromatidy sa rozchádzajú k pólom bunky, natiahnuté mikrotubulmi. Distribúcia chromatidov musí byť úplne rovnaká, pretože práve tento proces zabezpečuje udržanie konštantného počtu chromozómov v bunkách tela.
Na pódiu telofázy dcérske chromozómy sa zhromažďujú na póloch, despirujú, z vezikúl sa okolo nich vytvárajú jadrové membrány a v novovzniknutých jadrách sa objavujú jadierka.
Po jadrovom delení nastáva cytoplazmatické delenie - cytokinéza, pri ktorej dochádza k viac-menej rovnomernej distribúcii všetkých organel materskej bunky.
V dôsledku mitózy sa teda z jednej materskej bunky vytvoria dve dcérske bunky, z ktorých každá je genetickou kópiou materskej bunky (2n2c).
V chorých, poškodených, starnúcich bunkách a špecializovaných tkanivách tela môže nastať trochu iný proces delenia – amitóza. Amitóza nazývané priame delenie eukaryotických buniek, pri ktorom nedochádza k tvorbe geneticky ekvivalentných buniek, keďže bunkové zložky sú rozložené nerovnomerne. Nachádza sa v rastlinách v endosperme a u zvierat - v pečeni, chrupavke a rohovke oka.
meióza. Fázy meiózy
meióza je metóda nepriameho delenia primárnych zárodočných buniek (2n2c), ktorého výsledkom je vznik haploidných buniek (1n1c), najčastejšie zárodočných buniek.
Na rozdiel od mitózy pozostáva meióza z dvoch po sebe nasledujúcich bunkových delení, z ktorých každému predchádza interfáza. Prvé delenie meiózy (meióza I) sa nazýva redukcionista, pretože v tomto prípade je počet chromozómov polovičný a druhé delenie (meióza II) - rovnicové, keďže pri jeho procese sa zachováva počet chromozómov.

Medzifáza I prebieha ako medzifáza mitózy. Meióza I je rozdelená do štyroch fáz: profáza I, metafáza I, anafáza I a telofáza I. B profáza I Vyskytujú sa dva dôležité procesy: konjugácia a kríženie. Konjugácia- Ide o proces fúzie homológnych (párových) chromozómov po celej dĺžke. Páry chromozómov vytvorené počas konjugácie sú zachované až do konca metafázy I.
Prejsť- vzájomná výmena homológnych oblastí homológnych chromozómov. V dôsledku kríženia získajú chromozómy prijaté telom od oboch rodičov nové kombinácie génov, čo spôsobuje výskyt geneticky rôznorodých potomkov. Na konci profázy I, ako pri profáze mitózy, jadierko zmizne, centrioly sa rozchádzajú k pólom bunky a jadrová membrána sa rozpadne.

IN metafáza I páry chromozómov sú zarovnané pozdĺž rovníka bunky a k ich centromérom sú pripojené vretienkové mikrotubuly.
IN anafáza I Celé homológne chromozómy pozostávajúce z dvoch chromatidov sa rozchádzajú k pólom.
IN telofáza I Okolo zhlukov chromozómov na póloch bunky sa vytvárajú jadrové membrány a vytvárajú sa jadierka.
Cytokinéza I zabezpečuje separáciu cytoplazmy dcérskych buniek.
Dcérske bunky (1n2c) vytvorené ako výsledok meiózy I sú geneticky heterogénne, pretože ich chromozómy, náhodne rozptýlené na póloch buniek, obsahujú rôzne gény.
Porovnávacie charakteristiky mitózy a meiózy
| Podpísať | Mitóza | meióza | |
| Ktoré bunky sa začínajú deliť? | somatické (2n) | Primárne zárodočné bunky (2n) | |
| Počet divízií | 1 | 2 | |
| Koľko a aké bunky sa tvoria pri delení? | 2 somatické (2n) | 4 sexuálne (n) | |
| Medzifáza | Príprava bunky na delenie, zdvojnásobenie DNA | Veľmi krátke, nedochádza k zdvojeniu DNA | |
| Fázy | Meióza I | Meióza II | |
| Profáza | Môže dôjsť ku kondenzácii chromozómov, vymiznutiu jadierka, rozpadu jadrovej membrány, konjugácii a kríženiu | Kondenzácia chromozómov, vymiznutie jadierka, rozpad jadrovej membrány | |
| Metafáza | Páry chromozómov sú umiestnené pozdĺž rovníka, tvorí sa vreteno | Chromozómy sa zoradia pozdĺž rovníka, vytvorí sa vreteno | |
| Anaphase | Homologické chromozómy z dvoch chromatidov sa pohybujú smerom k pólom | Chromatidy sa pohybujú smerom k pólom | |
| Telofáza | Chromozómy despirujú, tvoria sa nové jadrové membrány a jadierka | Chromozómy despirujú, tvoria sa nové jadrové membrány a jadierka | |
Medzifáza II veľmi krátka, keďže v nej nedochádza k zdvojovaniu DNA, čiže neexistuje žiadna S-perióda.
Meióza II tiež rozdelené do štyroch fáz: profáza II, metafáza II, anafáza II a telofáza II. IN profáza II prebiehajú rovnaké procesy ako v profáze I, s výnimkou konjugácie a kríženia.
IN metafáza II chromozómy sú umiestnené pozdĺž rovníka bunky.
IN anafáza II chromozómy sú rozdelené na centroméroch a chromatidy sú natiahnuté smerom k pólom.
IN telofáza II Jadrové membrány a jadierka sa tvoria okolo zhlukov dcérskych chromozómov.
Po cytokinéza II Genetický vzorec všetkých štyroch dcérskych buniek je 1n1c, ale všetky majú odlišnú sadu génov, ktorá je výsledkom kríženia a náhodnej kombinácie chromozómov materských a otcovských organizmov v dcérskych bunkách.
Vývoj zárodočných buniek u rastlín a živočíchov
Gametogenéza(z gréčtiny gaméta- manželka, gaméty- manžel a genéza- vznik, vznik) je proces tvorby zrelých zárodočných buniek.
Keďže sexuálna reprodukcia si najčastejšie vyžaduje dvoch jedincov - samicu a muža, ktorí produkujú rôzne pohlavné bunky - vajíčka a spermie, potom musia byť procesy tvorby týchto gamét odlišné.
Povaha procesu do značnej miery závisí od toho, či sa vyskytuje v rastlinnej alebo živočíšnej bunke, pretože v rastlinách dochádza iba k mitóze pri tvorbe gamét a u zvierat dochádza k mitóze aj meióze.
Vývoj zárodočných buniek v rastlinách. U krytosemenných rastlín dochádza k tvorbe samčích a samičích reprodukčných buniek v rôznych častiach kvetu - tyčinkách a piestikoch.
Pred vytvorením mužských reprodukčných buniek - mikrogametogenéza(z gréčtiny mikr- malý) - stane sa mikrosporogenéza, teda tvorbu mikrospór v prašníkoch tyčiniek. Tento proces je spojený s meiotickým delením materskej bunky, ktorého výsledkom sú štyri haploidné mikrospóry. Mikrogametogenéza je spojená s mitotickým delením mikrospóry, pričom samčí gametofyt z dvoch buniek - veľký vegetatívny(sifonogénne) a plytké generatívny. Po rozdelení sa samčí gametofyt pokryje hustými membránami a vytvorí peľové zrno. V niektorých prípadoch sa generatívna bunka mitoticky delí aj počas procesu dozrievania peľu a niekedy až po prenesení na bliznu piestika a vytvoria sa dve nepohyblivé samčie zárodočné bunky - spermie. Po opelení sa z vegetatívnej bunky vytvorí peľová trubica, cez ktorú prenikajú spermie do vaječníka piestika na oplodnenie.

Vývoj samičích zárodočných buniek v rastlinách je tzv megagametogenéza(z gréčtiny megas- veľký). Vyskytuje sa vo vaječníku piestika, ktorému predchádza o megasporogenéza, v dôsledku čoho sa z materskej bunky megaspóry ležiacej v jadre meiotickým delením vytvoria štyri megaspóry. Jedna z megaspór sa mitoticky delí trikrát, čím sa získa ženský gametofyt – embryonálny vak s ôsmimi jadrami. Následným oddelením cytoplazmy dcérskych buniek sa z jednej z výsledných buniek stane vajíčko, po stranách ktorého ležia takzvané synergidy, na opačnom konci embryového vaku sa vytvoria tri antipódy a v strede , v dôsledku splynutia dvoch haploidných jadier vzniká diploidná centrálna bunka.
Vývoj zárodočných buniek u zvierat. U zvierat existujú dva procesy tvorby zárodočných buniek - spermatogenéza a oogenéza.
Spermatogenéza(z gréčtiny spermie, spermie- osivo a genéza- vznik, výskyt) je proces tvorby zrelých mužských zárodočných buniek - spermií. U ľudí sa vyskytuje v semenníkoch alebo semenníkoch a delí sa na štyri obdobia: rozmnožovanie, rast, dozrievanie a formovanie.

IN obdobie rozmnožovania primordiálne zárodočné bunky sa delia mitoticky, výsledkom čoho je vznik diploidov spermatogónie. IN obdobie rastu spermatogónie akumulujú živiny v cytoplazme, zväčšujú sa a menia sa na primárne spermatocyty, alebo spermatocyty 1. rádu. Až potom vstúpia do meiózy ( obdobie dozrievania), v dôsledku čoho vznikajú prvé dve sekundárny spermatocyt, alebo spermatocyt 2. rádu a potom - štyri haploidné bunky so stále dosť veľkým množstvom cytoplazmy - spermatidy. IN formačné obdobie stratia takmer všetku cytoplazmu a vytvoria bičík, ktorý sa zmení na spermie.
Spermie, alebo živí, - veľmi malé mobilné samčie reprodukčné bunky s hlavou, krkom a chvostom.

IN hlavu, okrem jadra, je akrozóm- modifikovaný Golgiho komplex, ktorý zabezpečuje rozpustenie vajíčok pri oplodnení. IN krčka maternice sú centrioly bunkového centra a základne konský chvost tvoria mikrotubuly, ktoré priamo podporujú pohyb spermií. Obsahuje tiež mitochondrie, ktoré poskytujú spermiám energiu ATP na pohyb.
Oogenéza(z gréčtiny OSN- vajcia a genéza- vznik, výskyt) je proces tvorby zrelých samičích zárodočných buniek - vajíčok. U ľudí sa vyskytuje vo vaječníkoch a pozostáva z troch období: rozmnožovania, rastu a dozrievania. Obdobia reprodukcie a rastu, podobné ako pri spermatogenéze, sa vyskytujú počas vnútromaternicového vývoja. V tomto prípade sa diploidné bunky tvoria z primárnych zárodočných buniek v dôsledku mitózy. oogónia, ktoré sa potom menia na diploidné primárne oocyty, alebo oocyty 1. rádu. Meióza a následná cytokinéza vyskytujúca sa v obdobie dozrievania, sa vyznačujú nerovnomerným delením cytoplazmy materskej bunky, takže v dôsledku toho sa najskôr získa sekundárny oocyt, alebo Oocyt 2. rádu, A prvé polárne teleso, a potom zo sekundárneho oocytu - vajíčka, ktoré si zachováva celú zásobu živín, a druhého polárneho telieska, pričom prvé polárne teliesko je rozdelené na dve časti. Polárne telesá pohlcujú nadbytočný genetický materiál.
U ľudí sa vajcia produkujú s intervalom 28-29 dní. Cyklus spojený s dozrievaním a uvoľňovaním vajíčok sa nazýva menštruačný.
Vajcia- veľká ženská reprodukčná bunka, ktorá nesie nielen haploidnú sadu chromozómov, ale aj významný prísun živín pre následný vývoj embrya.

Vajíčko u cicavcov je pokryté štyrmi membránami, ktoré znižujú pravdepodobnosť poškodenia rôznymi faktormi. Priemer vajíčka u ľudí dosahuje 150-200 mikrónov, zatiaľ čo u pštrosa to môže byť niekoľko centimetrov.
Bunkové delenie je základom pre rast, vývoj a rozmnožovanie organizmov. Úloha mitózy a meiózy
Ak v jednobunkových organizmoch vedie delenie buniek k zvýšeniu počtu jedincov, t. j. k reprodukcii, potom v mnohobunkových organizmoch môže mať tento proces rôzne významy. Rozdelenie embryonálnych buniek počnúc zygotou je teda biologickým základom vzájomne prepojených procesov rastu a vývoja. Podobné zmeny pozorujeme u ľudí v období dospievania, kedy sa počet buniek nielen zvyšuje, ale dochádza aj ku kvalitatívnej zmene v organizme. Rozmnožovanie mnohobunkových organizmov je založené aj na delení buniek, napríklad pri nepohlavnom rozmnožovaní sa vďaka tomuto procesu obnovuje celá časť organizmu a pri pohlavnom rozmnožovaní v procese gametogenézy vznikajú pohlavné bunky, ktoré následne viesť k vzniku nového organizmu. Je potrebné poznamenať, že hlavné metódy delenia eukaryotickej bunky - mitóza a meióza - majú v životných cykloch organizmov rôzny význam.
V dôsledku mitózy dochádza k rovnomernému rozdeleniu dedičného materiálu medzi dcérske bunky – presné kópie matky. Bez mitózy by existencia a rast mnohobunkových organizmov vyvíjajúcich sa z jedinej bunky, zygoty, bola nemožná, pretože všetky bunky takýchto organizmov musia obsahovať rovnakú genetickú informáciu.
Počas procesu delenia sa dcérske bunky stávajú čoraz rozmanitejšími v štruktúre a funkciách, čo súvisí s aktiváciou stále nových a nových skupín génov v nich v dôsledku medzibunkovej interakcie. Mitóza je teda nevyhnutná pre vývoj organizmu.
Tento spôsob delenia buniek je nevyhnutný pre procesy nepohlavného rozmnožovania a regenerácie (obnovy) poškodených tkanív, ako aj orgánov.
Meióza zase zabezpečuje stálosť karyotypu počas sexuálneho rozmnožovania, pretože pred pohlavným rozmnožovaním rozdeľuje sadu chromozómov na polovicu, ktorá sa potom obnoví v dôsledku oplodnenia. Okrem toho meióza vedie k vzniku nových kombinácií rodičovských génov v dôsledku kríženia a náhodnej kombinácie chromozómov v dcérskych bunkách. Vďaka tomu sa potomstvo ukazuje ako geneticky rôznorodé, čo poskytuje materiál pre prirodzený výber a je materiálnym základom pre evolúciu. Zmena počtu, tvaru a veľkosti chromozómov môže na jednej strane viesť k vzniku rôznych odchýlok vo vývoji organizmu až k jeho smrti a na druhej strane môže viesť k objaveniu sa jedincov viac prispôsobené prostrediu.
Bunka je teda jednotkou rastu, vývoja a reprodukcie organizmov.
RIEŠENIE PROBLÉMOV CYTOLÓGIE
Výchovno-metodická príručka
Nové Adelakovo 2014
Zostavil: učiteľ biológie L.I. Denisovej
Riešenie problémov v cytológii. Pre študentov ročníkov 9-11 / GBOU strednej školy v obci Novoe Adeljakovo“; komp. L.I. Denisová. – Nové Adelakovo, 2014.
Vzdelávacia a metodická príručka bola zostavená pre potreby učiteľov a študentov biológie pri príprave na záverečnú atestáciu, ako aj na biologické olympiády. Príručka obsahuje stručný teoretický materiál ku každému typu problému a príklady riešenia problémov. Určené pre študentov 9. – 11. ročníka inštitúcií všeobecného vzdelávania a učiteľov biológie.
Úvod …………………………………………………………………..
2.1 Ciele súvisí s určením percenta nukleotidov v DNA
2.2. R
2.3. Úlohy týkajúce sa konštrukcie molekuly mRNA, antikodónov tRNA a sekvencií aminokyselín v polypeptidovom reťazci. Job
Literatúra.
Úvod.
Riešenie problémov z cytológie je zahrnuté v KIM z biológie v Jednotnej štátnej skúške (úloha 39). Dokončenie tejto úlohy si vyžaduje podrobnú odpoveď a je zamerané na testovanie zručností
- aplikovať vedomosti v novej situácii;
- vytvoriť vzťahy medzi príčinami a následkami;
- analyzovať, systematizovať a integrovať poznatky;
- zovšeobecňovať a formulovať závery.
Na základe výsledkov analýzy výsledkov Jednotnej štátnej skúšky z biológie patria medzi slabo rozvinuté vedomosti a zručnosti účastníkov skúšky:
- stanovenie chromozómového súboru buniek vo vývojových cykloch rastlín;
- stanovenie počtu chromozómov a DNA v bunkách v rôznych fázach mitózy a meiózy;
- vysvetlenie a zdôvodnenie získaného výsledku.
Pri štúdiu biológie na základnej úrovni nie je dostatok času na praktické precvičovanie riešenia problémov v cytológii. Po absolvovaní príslušných tém, bez neustáleho opakovania praktickej praxe pri riešení problémov, sa zručnosti rýchlo zabúdajú. Študenti môžu mať túto príručku vždy po ruke, aby si zapamätali proces riešenia typických problémov. Navyše vo vidieckych oblastiach nie každý má voľný prístup k internetovým zdrojom.
Cytologické úlohy, ktoré sa objavujú v jednotnej štátnej skúške, možno rozdeliť do niekoľkých hlavných typov. Táto príručka ponúka riešenia problémov rôznych typov a poskytuje príklady pre samostatnú prácu. V prílohe je uvedená tabuľka genetického kódu použitého v riešení.
Táto vzdelávacia a metodická príručka bola zostavená s cieľom pomôcť učiteľom biológie študujúcim v 9. – 11. ročníku na stredných školách.
1.1 Kľúčové body, ktoré treba pamätať pri riešení problémov s cytológiou.
- Každá aminokyselina je dodávaná do ribozómov jednou tRNA, pretopočet aminokyselín v proteíne sa rovná počtu molekúl tRNApodieľa sa na syntéze bielkovín;
- Každá aminokyselina je kódovaná tromi nukleotidmi (jeden triplet alebo kodón), takže počet kódujúcich nukleotidov je vždy trikrát väčší apočet tripletov (kodónov) sa rovná počtu aminokyselín v proteíne;
- každá tRNA má antikodón komplementárny ku kodónu mRNA, tzvpočet antikodónov, a teda vo všeobecnosti molekuly tRNA rovný počtu mRNA kodónov;
- mRNA je komplementárna k jednému z reťazcov DNA, tzvpočet nukleotidov v mRNA sa rovná počtu nukleotidov v DNA. Počet trojčiat bude, samozrejme, tiež rovnaký.
Pri riešení množstva problémov v tejto časti je potrebné použiť tabuľku genetického kódu. Pravidlá používania tabuľky sú zvyčajne uvedené v zadaní, ale je lepšie sa to naučiť vopred. Ak chcete určiť aminokyselinu kódovanú konkrétnym tripletom, musíte vykonať nasledujúce kroky:
- prvý nukleotid tripletu sa nachádza v ľavom zvislom rade,
- druhý - v hornej horizontále,
- tretí je v pravom vertikálnom rade.
- aminokyselina zodpovedajúca tripletu sa nachádza v bode priesečníka imaginárnych čiar, pochádzajúce z nukleotidov.
1.2. Písomná dokumentácia riešenia problému.
odpoveď:
Dôležitým bodom pri riešení úloh je vysvetliť vykonávané činnosti, najmä ak úloha hovorí: „Vysvetlite svoju odpoveď. Prítomnosť vysvetlení umožňuje hodnotiteľovi vyvodiť záver o tom, ako študent rozumie tejto téme, a ich absencia môže viesť k strate veľmi dôležitého bodu. Úloha 39 má hodnotu troch bodov, ktoré sa udeľujú v prípade úplne správneho riešenia. Preto pri spustení úlohy je v prvom rade potrebné zdôrazniť všetky problémy. Počet odpovedí im musí zodpovedať.
Kapitola 2. Typy úloh v cytológii.
2.1 Ciele súvisí s určením percenta nukleotidov v DNA
Ešte pred objavom Watsona a Cricka, v roku 1950, austrálsky biochemik Edwin Chargaff zistil, žeže v DNA akéhokoľvek organizmu sa počet adenylových nukleotidov rovná počtu tymidylových nukleotidov a počet guanylových nukleotidov sa rovná počtu cytosylových nukleotidov (A=T, G=C) alebo celkovému počtu purínových dusíkatých zásad sa rovná celkovému počtu pyrimidínových dusíkatých zásad (A+G=C+T ).Tieto vzory sa nazývajú „Chargaffove pravidlá“.
Faktom je, že keď sa vytvorí dvojitá špirála, dusíkatá báza tymín je vždy inštalovaná oproti dusíkatej báze adenínu v jednom reťazci a cytozín je inštalovaný oproti guanínu, to znamená, že reťazce DNA sa navzájom dopĺňajú. A tieto párové nukleotidy sú navzájom komplementárne (z latinského komplementum – sčítanie).
Prečo sa tento princíp dodržiava? Na zodpovedanie tejto otázky si musíme pamätať chemickú povahu dusíkatých heterocyklických zásad. Adenín a guanín patria k purínom a cytozín a tymín patria k pyrimidínom, to znamená, že medzi dusíkatými zásadami rovnakej povahy nie sú vytvorené väzby. Navyše komplementárne bázy si navzájom zodpovedajú geometricky, t.j. vo veľkosti a tvare. Nukleotidová komplementarita je teda chemická a geometrická zhoda štruktúr ich molekúl navzájom.
Dusíkaté bázy obsahujú vysoko elektronegatívne atómy kyslíka a dusíka, ktoré nesú čiastočne záporný náboj, ako aj atómy vodíka, ktoré nesú čiastočne kladný náboj. Vďaka týmto čiastkovým nábojom vznikajú vodíkové väzby medzi dusíkatými bázami antiparalelných sekvencií molekuly DNA.
Úloha. V molekule DNA je 23 % adenylnukleotidov z celkového počtu nukleotidov. Určte počet tymidylových a cytosylových nukleotidov.
Odpoveď: T=23 %; C=27 %
Úloha Uvádza sa molekula DNA s relatívnou molekulovou hmotnosťou 69 tisíc, z toho 8625 adenylnukleotidov. Relatívna molekulová hmotnosť jedného nukleotidu je v priemere 345. Koľko jednotlivých nukleotidov je v tejto DNA? Aká je dĺžka jeho molekuly?
Vzhľadom na to: M(r) DNA – 69000 Množstvo A – 8625 M(r) nukleotidu – 345 Definuj: Počet nukleotidov v DNA | Riešenie : 1. Určte, koľko adenylových nukleotidov je v danej molekule DNA: 8625: 345 = 25. 2. Podľa Chargaffovho pravidla platí A = G, t.j. v danej molekule DNA A=T=25. 3. Určte, koľko z celkovej molekulovej hmotnosti tejto DNA tvorí podiel guanylových nukleotidov: 69 000 – (8625x2) = 51 750. 4. Určte celkový počet guanylových a cytosylových nukleotidov v tejto DNA: 51 750:345=150. 5. Oddelene stanovte obsah guanylových a cytosylových nukleotidov: 150:2 = 75; 6. Určte dĺžku tejto molekuly DNA: (25 + 75) x 0,34 = 34 nm. |
Odpoveď : A = T = 25; G=C=75; 34 nm.
2.2. R Výpočtové úlohy venované určovaniu počtu aminokyselín v proteíne, ako aj počtu nukleotidov a tripletov v DNA alebo RNA.
- Aminokyseliny potrebné na syntézu proteínov sú dodávané do ribozómov pomocou tRNA. Každá molekula tRNA nesie iba jednu aminokyselinu.
- Informácie o primárnej štruktúre molekuly proteínu sú zakódované v molekule DNA.
- Každá aminokyselina je kódovaná sekvenciou troch nukleotidov. Táto sekvencia sa nazýva triplet alebo kodón.
Úloha: Na translácii sa podieľalo 30 molekúl tRNA. Určte počet aminokyselín, ktoré tvoria výsledný proteín, ako aj počet tripletov a nukleotidov v géne, ktorý tento proteín kóduje.
odpoveď: Počet aminokyselín – 30. Počet tripletov – 30. Počet nukleotidov – 90.
Úloha: Podľa niektorých vedcov je celková dĺžka všetkých molekúl DNA v jadre jednej ľudskej zárodočnej bunky asi 102 cm Koľko nukleotidových párov obsahuje DNA jednej bunky (1 nm = 10–6 mm)?
Odpoveď: 3x109 párov.
2.3. Úlohy týkajúce sa konštrukcie molekuly mRNA, antikodónov tRNA a sekvencií aminokyselín v polypeptidovom reťazci. Jobs tabuľkou genetického kódu.
Úloha: RNA obsahuje uracil namiesto tymínu. Biosyntéza proteínov zahŕňala t-RNA s antikodónmi: UUA, GGC, CGC, AUU, CGU. Určite nukleotidovú sekvenciu úseku každého reťazca molekuly DNA, ktorá nesie informácie o syntetizovanom polypeptide, a počet nukleotidov obsahujúcich adenín, guanín, tymín, cytozín v molekule dvojvláknovej DNA
Vzhľadom na to: T-RNA - UUA, GGC, CGC, AUU, TsGU | Riešenie:
Číslo A=T=7, číslo G=C=8 |
Definuj: Nukleotidová sekvencia sekcie každého vlákna molekuly DNA Počet nukleotidov obsahujúcich adenín, guanín, tymín, cytozín v molekule DNA |
Úloha: fragment jedného z reťazcov DNA má nasledujúcu štruktúru: AAGGCTACTGTTG. Postavte na ňom mRNA a určte poradie aminokyselín vo fragmente molekuly proteínu.
Odpoveď: fen-arg-cis-asn.
Úloha: Ribozómy z rôznych buniek, celý súbor aminokyselín a identické molekuly mRNA a tRNA boli umiestnené do skúmavky a boli vytvorené všetky podmienky pre syntézu proteínov. Prečo sa v skúmavke bude jeden typ proteínu syntetizovať na rôznych ribozómoch.
odpoveď: Rovnaký proteín sa syntetizuje na rovnakej mRNA, pretože informácie sú rovnaké.
- Úloha: Fragment mRNA má nasledujúcu štruktúru: GAUGAGUATSUUCAA. Určite tRNA antikodóny a aminokyselinovú sekvenciu kódovanú v tomto fragmente. Napíšte aj fragment molekuly DNA, na ktorej bola táto mRNA syntetizovaná.
Odpoveď : Sekvencia aminokyselín - : asp-glu-tyr-fen-lys. T-RNA antikodóny - TsUA, TsUC, AUG, AAG, UUU. Fragment DNA – CTATTSCATGAAGTTT
2.4.Úlohy na určenie počtu molekúl DNA a chromozómov v procese mitózy a meiózy.
Mitóza - hlavná metóda delenia eukaryotických buniek, pri ktorej najskôr dochádza k zdvojeniu a potom je dedičný materiál rovnomerne rozdelený medzi dcérske bunky.
Mitóza je nepretržitý proces so štyrmi fázami: profáza, metafáza, anafáza a telofáza. Pred mitózou sa bunka pripravuje na delenie alebo interfázu. Obdobie prípravy bunky na mitózu a samotná mitóza tvoria spolumitotický cyklus.
Medzifáza pozostáva z troch období: presyntetické alebo postmitotické, - G 1 , syntetické - S, postsyntetické alebo premitotické - G 2 .
Predsyntetické obdobie(2 n 2 c , kde n - počet chromozómov, s - počet molekúl DNA) - rast buniek, aktivácia procesov biologickej syntézy, príprava na ďalšie obdobie.
Syntetické obdobie(2 n 4 c ) - replikácia DNA.
POZOR!
Po zdvojení zostáva sada chromozómov diploidná (2n), pretože sesterské chromatidy zostávajú spojené na centromére.
Postsyntetické obdobie(2 n 4 c ) - príprava bunky na mitózu, syntézu a akumuláciu bielkovín a energie pre nadchádzajúce delenie, zvýšenie počtu organel, zdvojnásobenie centriolov.
Profáza (2 n 4 c ) - demontáž jadrových membrán, divergencia centriolov k rôznym pólom bunky, tvorba vretenových filamentov, „miznutie“ jadier, kondenzácia bichromatidových chromozómov.
Metafáza (2 n 4 s ) - zarovnanie maximálne kondenzovaných bichromatidových chromozómov v ekvatoriálnej rovine bunky (metafázová platnička), prichytenie závitov vretienka na jednom konci k centriolom, druhému k centromérom chromozómov.
Anafáza (4 n 4 c ) - rozdelenie dvojchromatidových chromozómov na chromatidy a divergencia týchto sesterských chromatidov k opačným pólom bunky (v tomto prípadechromatidy sa stávajú samostatnými jednochromatidovými chromozómami).
Telofáza (2 n 2 c v každej dcérskej bunke) - dekondenzácia chromozómov, tvorba jadrových membrán okolo každej skupiny chromozómov, rozpad vretienkových závitov, objavenie sa jadierka, delenie cytoplazmy (cytotómia). Cytotómia v živočíšnych bunkách sa vyskytuje v dôsledku štiepnej brázdy, v rastlinných bunkách - v dôsledku bunkovej dosky.
meióza - ide o špeciálnu metódu delenia eukaryotických buniek, v dôsledku ktorej bunky prechádzajú z diploidného stavu do haploidného stavu. Meióza pozostáva z dvoch po sebe nasledujúcich delení, ktorým predchádza jediná replikácia DNA.
Prvé meiotické delenie (meióza 1)sa nazýva redukcia, pretože práve pri tomto delení sa počet chromozómov zníži na polovicu: z jednej diploidnej bunky (2 n 4 s ) dva haploidy (1 n2c).
Medzifáza 1 (na začiatku - 2 n 2 s, na konci - 2 n 4 s ) - syntéza a akumulácia látok a energie potrebných pre obe delenia, zväčšenie veľkosti buniek a počtu organel, zdvojnásobenie centriol, replikácia DNA, ktorá končí profázou 1.
Profáza 1 (2 n 4 s ) - demontáž jadrových membrán, divergencia centriolov k rôznym pólom bunky, tvorba vretenových filamentov, „miznutie“ jadierok, kondenzácia bichromatidových chromozómov, konjugácia homológnych chromozómov a kríženie.
Metafáza 1 (2 n 4 s ) - zoradenie bivalentov v ekvatoriálnej rovine bunky, pripojenie vretenových filamentov na jednom konci k centriolám, na druhom k centromérom chromozómov.
Anafáza 1 (2 n 4 s ) - náhodné nezávislésegregácia dichromatidových chromozómovna opačné póly bunky (z každého páru homológnych chromozómov ide jeden chromozóm na jeden pól, druhý na druhý), rekombinácia chromozómov.
Telofáza 1 (1 n 2 c v každej bunke) - tvorba jadrových membrán okolo skupín dichromatidových chromozómov, delenie cytoplazmy. V mnohých rastlinách bunka prechádza z anafázy 1 okamžite do profázy 2.
Druhé meiotické delenie (meióza 2) nazývané rovnicové.
Interfáza 2 alebo interkinéza (1n 2c ), je krátka prestávka medzi prvým a druhým meiotickým delením, počas ktorej nedochádza k replikácii DNA. Charakteristické pre živočíšne bunky.
Profáza 2 (1 n 2 c ) - demontáž jadrových membrán, divergencia centriolov k rôznym pólom bunky, tvorba vretenových filamentov.
Metafáza 2 (1 n 2 c ) - zarovnanie bichromatidových chromozómov v ekvatoriálnej rovine bunky (metafázová platnička), prichytenie vretenových filamentov jedným koncom k centriolám, druhým k centromérom chromozómov; 2 blok oogenézy u ľudí.
Anafáza 2 (2 n 2 s ) - rozdelenie dvojchromatidových chromozómov na chromatidy a divergencia týchto sesterských chromatidov na opačné póly bunky (v tomto prípade sa chromatidy stávajú samostatnými jednochromatidovými chromozómami), rekombinácia chromozómov.
Telofáza 2 (1 n 1 c v každej bunke) - dekondenzácia chromozómov, tvorba jadrových membrán okolo každej skupiny chromozómov, rozpad vlákien vretienka, objavenie sa jadierka, delenie cytoplazmy (cytotómia) s výsledným vytvorením štyroch haploidných buniek.

Úloha: Hovädzí dobytok má v somatických bunkách 60 chromozómov. Určte počet chromozómov a molekúl DNA v bunkách vaječníkov v interfáze pred začiatkom delenia a po delení meiózou I. Vysvetlite, ako vzniká taký počet chromozómov a molekúl DNA.
odpoveď: V interfáze je 60 chromozómov a 120 molekúl DNA. Po meióze I je 30 chromozómov a 60 molekúl DNA.
Úloha: Chromozómový súbor somatických buniek pšenice je 28. Určte chromozómový súbor a počet molekúl DNA v jadre (bunke) vajíčka pred začiatkom meiózy I a meiózy II. Vysvetlite výsledky v každom prípade.
odpoveď: Pred začiatkom meiózy I = 28 chromozómov, 56 molekúl DNA. Pred meiózou II = 14 chromozómov, 28 molekúl DNA
Úloha: Je známe, že somatické bunky kapusty obsahujú 18 chromozómov. Stanovte chromozómovú sadu a počet molekúl DNA v jednej z buniek vajíčka pred začiatkom meiózy, v anafáze meiózy I a v anafáze meiózy II. Vysvetlite, aké procesy prebiehajú v týchto obdobiach a ako ovplyvňujú zmeny v počte DNA a chromozómov.
odpoveď: Pred začiatkom meiózy - 18 chromozómov, 36 molekúl DNA. V anafáze meiózy I je 18 chromozómov, 36 molekúl DNA. V anafáze je 18 chromozómov, 18 molekúl DNA.
2.5. Úlohy určiť dĺžku samostatného úseku DNA alebo počet nukleotidov v ňom.
Podľa modelu amerického biochemika J. Watsona a anglického fyzika F. Cricka sú molekuly DNA dva pravotočivé polynukleotidové reťazce alebo dvojitá špirála, skrútené okolo spoločnej osi. Na jednu otáčku špirály pripadá približne 10 nukleotidových zvyškov. Reťaze v tejto dvojitej špirále sú antiparalelné, to znamená nasmerované v opačných smeroch, takže 3" koniec jednej reťaze je umiestnený oproti 5" koncu druhej. Veľkosti molekúl DNA sú zvyčajne vyjadrené počtom nukleotidov, ktoré ich tvoria. Tieto veľkosti sa líšia od niekoľkých tisíc nukleotidových párov v bakteriálnych plazmidoch a niektorých vírusoch až po mnoho stoviek tisíc nukleotidových párov vo vyšších organizmoch.
Lineárna dĺžka jedného nukleotidu v nukleovej kyseline
Ln = 0,34 nm = 3,4 angstromov
Priemerná molekulová hmotnosť jedného nukleotidu
Mr n = 345 amu (da)
Úloha. Dĺžka obrysu molekuly bakteriofágovej DNA je 17x10" 6 Po vystavení mutagénom sa ukázalo, že dĺžka je 13,6 x 10-6 m) Určte, koľko párov dusíkatých báz sa stratilo v dôsledku mutácie, ak je známe, že vzdialenosť medzi susednými nukleotidmi je 34x10 11 m.
Vzhľadom na to: Dĺžka obrysu molekuly DNA - 17x10" 6 m Po expozícii - 13,6x10-6 m Vzdialenosť medzi susednými nukleotidmi je - 34x10 11 m. | Riešenie. 1) Vypočítajme celkovú dĺžku segmentu DNA bakteriofága strateného v dôsledku vystavenia mutagénom. 17x10"6 - 13,6x106 = 3,4x106(m). 2) Vypočítajme počet nukleotidových párov vo vypadnutom fragmente: 3,4x10-6 / 34x10"11 = 104 = 10 LLC (nukleotidových párov) |
Definuj: koľko párov dusíkových báz sa stratilo v dôsledku mutácie |
odpoveď: 10 tisíc párov báz.
Kapitola 4. Príklady úloh na samostatné riešenie.
- Molekula DNA obsahuje 31 % adenínu. Určte, koľko (v %) obsahuje táto molekula ďalších nukleotidov.
- Na translácii sa podieľalo 50 molekúl tRNA. Určte počet aminokyselín, ktoré tvoria výsledný proteín, ako aj počet tripletov a nukleotidov v géne, ktorý tento proteín kóduje.
- Fragment DNA pozostáva zo 72 nukleotidov. Určte počet tripletov a nukleotidov v mRNA, ako aj počet aminokyselín, ktoré tvoria výsledný proteín.
- Fragment jedného z reťazcov DNA má nasledujúcu štruktúru: GGCTCTAGCTTC. Postavte na ňom mRNA a určte poradie aminokyselín vo fragmente molekuly proteínu (použite na to tabuľku genetických kódov).
- Fragment mRNA má nasledujúcu štruktúru: GCUAAUGUUCUUUAC. Určite tRNA antikodóny a aminokyselinovú sekvenciu kódovanú v tomto fragmente. Napíšte aj fragment molekuly DNA, na ktorej bola táto mRNA syntetizovaná (použite na to tabuľku genetického kódu).
- Fragment DNA má nasledujúcu nukleotidovú sekvenciu AGCCGACTTGCC. Určte nukleotidovú sekvenciu tRNA, ktorá je syntetizovaná na tomto fragmente, a aminokyselinu, ktorú táto tRNA ponesie, ak tretí triplet zodpovedá antikodónu tRNA. Na vyriešenie úlohy použite tabuľku genetického kódu.
- V živočíšnej bunke je diploidná sada chromozómov 20. Určte počet molekúl DNA pred mitózou, po mitóze, po prvom a druhom delení meiózy.
- 15 molekúl glukózy vstúpilo do disimilácie. Určte množstvo ATP po glykolýze, po energetickej fáze a celkový efekt disimilácie.
- Do Krebsovho cyklu vstúpilo 6 molekúl PVC. Určte množstvo ATP po energetickej fáze, celkový účinok disimilácie a počet molekúl glukózy, ktoré vstúpili do disimilácie.
Odpovede:
- T = 31 %, G = C = 19 % každý.
- 50 aminokyselín, 50 tripletov, 150 nukleotidov.
- 24 tripletov, 24 aminokyselín, 24 molekúl tRNA.
- mRNA: CCG-AGA-UCG-AAG. Aminokyselinová sekvencia: pro-arg-ser-lys.
- DNA fragment: CGATTACAAGAAATG. T-RNA antikodóny: CGA, UUA, CAA, GAA, AUG. Aminokyselinová sekvencia: ala-asn-val-ley-tyr.
- tRNA: UCG-GCU-GAA-CHG. Antikodón GAA, kodón mRNA - CUU, prenesená aminokyselina - leu.
- 2n = 20. Genetická sada:
- pred mitózou 40 molekúl DNA;
- po mitóze 20 molekúl DNA;
- po prvom meiotickom delení 20 molekúl DNA;
- po druhom meiotickom delení je 10 molekúl DNA.
- Pretože jedna molekula glukózy produkuje 2 molekuly PVK a 2ATP, syntetizuje sa 30 ATP. Po energetickom štádiu disimilácie sa vytvorí 36 molekúl ATP (s rozpadom 1 molekuly glukózy), preto sa syntetizuje 540 ATP. Celkový efekt disimilácie je 540+30=570 ATP.
- Do Krebsovho cyklu vstúpilo 6 molekúl PVA, preto sa rozpadli 3 molekuly glukózy. Množstvo ATP po glykolýze je 6 molekúl, po energetickom štádiu - 108 molekúl, celkový efekt disimilácie je 114 molekúl ATP.
Literatúra:
Kontrolné úlohy z genetiky a metodické odporúčania na ich realizáciu. [komp. L.I. Lushina, S.V. Zalyashchev, A.A. Semenov, O. N. Nošková]. – Samara: SGPU, 2007, 142 s.
http://ege-study.ru
http://licey.net
http://reshuege.ru
http://www.fipi.ru
Príloha I Genetický kód (mRNA)
Prvá základňa | Druhá základňa | Tretia základňa |
|||
Fén | Ser | Strelecká galéria | Cis | ||
Fén | Ser | Strelecká galéria | Cis | ||
Lei | Ser | ||||
Lei | Ser | Tri | |||
Lei | O | Gies | Arg | ||
Lei | O | Gies | Arg | ||
Lei | O | Gln | Arg | ||
Lei | O | Gln | Arg | ||
Ile | Tre | Asn | Ser | ||
Ile | Tre | Asn | Ser | ||
Ile | Tre | Liz | Arg | ||
Meth | Tre | Liz | Arg | ||
Šachta | Ala | Asp | Gli | ||
Šachta | Ala | Asp | Gli | ||
Šachta | Ala | Glu | Gli | ||
Šachta | Ala | Glu | Gli |
V lekcii sa naučíme históriu vzniku cytológie, zapamätáme si pojem bunky a zvážime prínos rôznych vedcov k rozvoju cytológie.
Všetky živé bytosti, s výnimkou vi-ru-s, sú tvorené bunkami. Ale pre vedcov z minulosti nebola bunková štruktúra živých organizmov taká zrejmá ako pre vás a mňa. Veda, ktorá študuje bunku, cytológie, vznikla až v polovici 19. storočia. Bez znalosti toho, odkiaľ život pochádza, že sa objavuje v najmenšej jednotke, sa až do stredoveku objavovali teórie, že napríklad žaby pochádzajú zo špiny a myši sa rodia v špinavej spodnej bielizni (obr. 2).

Ryža. 2. Teórie stredoveku ()
„Špinavá bielizeň vedy stredného storočia“ bola prvýkrát „šitá“ v roku 1665. Anglická príroda-is-py-ta-tel Ro- Bert Hooke (obr. 3).

Ryža. 3. Robert Hooke ()
Prvýkrát si prezrel a opísal schránky rastlinných buniek. A už v roku 1674 bol pod vlastnoručne vyrobeným mikom ako prvý vidieť jeho holandský coll-le-ha An-to-ni van Leeuwen-hoek (obr. 4) -súbor niektorých jednoduchých a samostatných živých buniek, napr. ako erytrocyty a spermie -áno.

Ryža. 4. Anthony van Leeuwenhoek ()
Výskum Le-ven-gu-ka sa stal takým fan-ta-sti-che-ski-mi, že v roku 1676 Londýnska Co-ro-lion-society, kam poslal výsledky svojho výskumu, veľmi v nich pre-so-mnou. Existencia jednobunkových orgánov a krviniek napríklad nezapadá do rámca toho, kde je veda.
Pochopenie výsledkov práce holandského vedca trvalo niekoľko storočí. Až do polovice 19. stor. Nemecký vedec Theodor Schwann na základe práce svojho kolegu Ma-tti-a-sa Schlei-de-na (obr. 5) tvoria základné princípy bunkovej teórie, ktoré používame dodnes.

Ryža. 5. Theodor Schwann a Matthias Schleiden ()
Schwann si uvedomil, že bunky rastlín a živočíchov majú spoločný princíp štruktúry, pretože sa akým spôsobom tvoria rovnako; všetky bunky sú sebestačné a každý organizmus je súborom skupín buniek živých jedincov (obr. 6).

Ryža. 6. Červené krvinky, delenie buniek, molekula DNA ()
Ďalší výskum vedeckých pozícií vytvorí základné princípy modernej bunkovej teórie:
- Bunka je univerzálna štrukturálna jednotka života.
- Bunky sa množia delením (bunka od bunky).
- Bunky sú uložené, re-ra-ba-you-va-yut, re-a-li-zu-yut a re-y-y-yat na postupnú informačnú formu-ma -áciu.
- Bunka je od určitej štrukturálnej úrovne organizácie živej ma-te-rii najmocnejší bio-systém.
- Viacbunkové presné orga-niz-sme komplex vzájomne pôsobiacich systémov rôznych buniek, ktoré poskytujú chi-va-yu-shchih or-ga-low-mu rast, vývoj, metabolizmus a energiu.
- Bunky všetkých organizmov sú si navzájom podobné štruktúrou, zložením a funkciou.
Bunky sú rôzne. Môžu sa líšiť štruktúrou, tvarom a funkciou (obr. 7).

Ryža. 7. Bunková diverzita ()
Medzi nimi sú voľne žijúce bunky, ktoré sa správajú ako jednotlivci populácií a druhov, ako ich vlastné organizmy.ga-bottom-we. Ich vitalita nezávisí len od toho, ako fungujú vnútorné bunkové štruktúry, or-ga-ale-a-y. Sami si potrebujú zaobstarať potravu, pohybovať sa v prostredí, množiť sa, čiže pôsobiť ako malí, no celkom sebestační jedinci. Takých slobodných je veľa. Sú zahrnuté vo všetkých kráľovstvách bunkovej živej prírody a obývajú všetky prostredia života na našej planéte. Vo viacbunkovom presnom organ-ga-dne je bunka jeho súčasťou, z buniek sa tvoria tkanivá a orgán-ga -us.
Veľkosť buniek môže byť veľmi odlišná - od jednej desatiny mikrónu do 15 centimetrov - to je veľkosť vajíčka krajiny, ktorá predstavuje jednu bunku, a hmotnosť tejto bunky je pol kilogramu. A to nie je limit: napríklad vajcia di-no-saurov môžu dosahovať dĺžku až 45 centimetrov (obr. 8) .

Ryža. 8. Dinosaurie vajce ()
Vo viacbunkových organizáciách zvyčajne rôzne bunky vykonávajú rôzne funkcie. Bunky podobnej štruktúry, umiestnené v blízkosti, spojené medzibunkovou substanciou a účelom - potrebné na vykonávanie určitých funkcií v organizácii, tvoria tkanivá (obr. 9).

Ryža. 9. Tvorba tkaniva ()
Život je veľa or-ga-niz-ma v závislosti od toho, ako slabo fungujú bunky, ktoré vstupujú do svojho zloženia. Preto si bunky medzi sebou nekonkurujú, naopak, dochádza k spolupráci a špecializácii ich funkcií. Možné or-ga-niz-mu prežiť v tých si-tu-a-tsi-yah, v ktorých jednotlivé bunky nie si-li-v-va- ut. V zložitých mnohobunkových organizmoch - rastlinách, zvieratách a ľuďoch - bunky or-ga-ni- zón v tkanine, tkanivách - v orgánoch, orgánoch - v systéme orgánov. A každý z týchto systémov funguje tak, aby zabezpečil existenciu celej organizácie.
Napriek všetkým rôznym tvarom a veľkostiam sú bunky rôznych typov navzájom podobné. Procesy ako dýchanie, biosyntéza, metabolizmus prebiehajú v bunkách bez ohľadu na to, či sú -but-kle-toch-ny-mi alebo-ga-niz-ma-mi alebo sú zahrnuté v zložení mnohých-kle-presne- žiadne bytosti. Každá bunka prijíma potravu, čerpá z nej energiu a prijíma energiu z odpadových produktov.spoločnosti, ktorá si udržiava stabilitu vlastného hi-mi-che-s-co-sta-va a sama sa reprodukuje, teda vykonáva všetky procesy, od ktorých závisí jej život.
To všetko nám umožňuje považovať bunku za špeciálnu jednotku živej hmoty, za elementárny živý systém (obr. 10).

Ryža. 10. Schematický nákres bunky ()
Všetky živé tvory, od in-fu-zo-ria až po slona alebo veľrybu, najväčšieho dnešného cicavca wow, sú vyrobené z buniek. Jediný rozdiel je v tom, že in-fu-zo-rii sú najviac skladovateľné bio-systémy, ktoré pozostávajú z jednej bunky, a bunky veľryby sú or-ga-ni-zo-va-ny a inter-and-mo. -spojené ako časti veľkého 190-tonového celku. Stav celej or-ga-niz-ma závisí od toho, ako fungujú jej časti, teda bunky.
Bibliografia
- Mamontov S.G., Zakharov V.B., Agafonova I.B., Sonin N.I. Biológia. Všeobecné vzory. - Drop, 2009.
- Ponomareva I.N., Kornilova O.A., Chernova N.M. Základy všeobecnej biológie. 9. ročník: Učebnica pre žiakov 9. ročníka všeobecnovzdelávacích inštitúcií / Ed. Prednášal prof. I.N. Ponomareva. - 2. vyd., prepracované. - M.: Ventana-Graf, 2005
- Pasechnik V.V., Kamensky A.A., Kriksunov E.A. Biológia. Úvod do všeobecnej biológie a ekológie: Učebnica pre 9. ročník, 3. vyd., stereotyp. - M.: Drop, 2002.
- Krugosvet.ru ().
- Uznaem-kak.ru ().
- Mewo.ru ().
Domáca úloha
- Čo študuje cytológia?
- Aké sú hlavné ustanovenia bunkovej teórie?
- Ako sa líšia bunky?
Mišnina Lidija Alexandrovna
učiteľ biológie
MBOU stredná škola č. 3 Obec Akbulak
Trieda 11
Príprava na jednotnú štátnu skúšku: riešenie úloh z cytológie
V metodických odporúčaniach na skvalitnenie výučby biológie, vypracovaných na základe rozboru ťažkostí absolventov na Jednotnej štátnej skúške v roku 2014, autori G.S. Kalinová, R.A. Petrosovej je nízka úroveň dokončenia úloh na určenie počtu chromozómov a DNA v rôznych fázach mitózy alebo meiózy.
Úlohy v skutočnosti nie sú také ťažké, aby spôsobovali vážne ťažkosti. Čo treba brať do úvahy pri príprave absolventov na túto problematiku?
Riešenie cytologických problémov si vyžaduje nielen znalosti o mitóze a meióze, ich fázach a udalostiach v nich prebiehajúcich, ale aj povinné vlastníctvo informácií o štruktúre a funkciách chromozómov, množstve genetického materiálu v bunke.
Preto začíname našu prípravu preskúmaním materiálu o chromozómoch. Zameriavame sa na skutočnosť, že chromozómy sú nukleoproteínové štruktúry v jadre eukaryotickej bunky.
Obsahujú asi 99 % celkovej DNA bunky, zvyšok DNA sa nachádza v iných bunkových organelách, ktoré určujú cytoplazmatickú dedičnosť. DNA v chromozómoch eukaryotov je v komplexe s hlavnými proteínmi – histónmi a s nehistónovými proteínmi, ktoré zabezpečujú komplexné balenie DNA v chromozómoch a reguláciu jej schopnosti syntetizovať ribonukleové kyseliny (RNA) – transkripciu.
Vzhľad chromozómov sa výrazne mení v rôznych štádiách bunkového cyklu a ako kompaktné štruktúry s charakteristickou morfológiou sú chromozómy jasne rozlíšiteľné vo svetelnom mikroskope len v období delenia buniek.
V štádiu metafázy mitózy a meiózy sa chromozómy skladajú z dvoch pozdĺžnych kópií, ktoré sa nazývajú sesterské chromatidy a ktoré sa tvoria počas replikácie DNA počas S periódy interfázy. V metafázových chromozómoch sú sesterské chromatidy spojené v primárnom zúžení nazývanom centroméra. Centroméra je zodpovedná za oddelenie sesterských chromatidov do dcérskych buniek počas delenia
Kompletná sada chromozómov v bunke, charakteristická pre daný organizmus, sa nazýva karyotyp. V ktorejkoľvek bunke tela väčšiny zvierat a rastlín je každý chromozóm zastúpený dvakrát: jeden z nich je prijatý od otca, druhý od matky počas fúzie jadier zárodočných buniek počas procesu oplodnenia. Takéto chromozómy sa nazývajú homológne a súbor homológnych chromozómov sa nazýva diploidný.
Teraz si môžete zopakovať materiál o delení buniek.
Z medzifázových dejov považujeme len syntetické obdobie, aby sme nerozptyľovali pozornosť školákov, ale sústredili sa len na správanie sa chromozómov.
Pripomeňme si: počas syntetického (S) obdobia dochádza k zdvojnásobeniu genetického materiálu prostredníctvom replikácie DNA. Dochádza k nej polokonzervatívnym spôsobom, keď sa dvojzávitnica molekuly DNA rozchádza na dva reťazce a na každom z nich sa syntetizuje komplementárny reťazec.
Výsledkom sú dva identické dvojité závitnice DNA, z ktorých každý pozostáva z jedného nového a jedného starého vlákna DNA. Množstvo dedičného materiálu sa zdvojnásobí, ale počet chromozómov zostáva rovnaký – chromozóm sa stáva bichromatidným (2n4c).
Zvážte správanie chromozómov počas mitózy:
- V profáze, metafáze - 2p 4s - keďže nedochádza k deleniu buniek;
- Počas anafázy sa chromatidy oddelia, počet chromozómov sa zdvojnásobí (chromatidy sa stanú samostatnými chromozómami, ale zatiaľ sú všetky v jednej bunke) 4n 4c;
- v telofáze 2n2c (jednochromatidové chromozómy zostávajú v bunkách).
Opakujte meiózu:
- V profáze 1, metafáze 1, anafáze 1 - 2p 4s - keďže nedochádza k deleniu buniek;
- v telofáze - zostáva p2c, keďže po divergencii homológnych chromozómov zostáva v bunkách haploidná sada, ale chromozómy sú bichromatidné;
- V profáze 2, metafáze 2, ako aj telofáze 1 - p2s;
- Venujte zvláštnu pozornosť anafáze 2, pretože po divergencii chromatíd sa počet chromozómov zvýši 2-krát (chromatidy sa stanú nezávislými chromozómami, ale zatiaľ sú všetky v jednej bunke) 2n 2c;
- v telofáze 2 - ps (jednochromatidové chromozómy zostávajú v bunkách.
Až teraz, keď sú deti teoreticky pripravené, môžeme prejsť k riešeniu problémov.
Typická chyba pri príprave maturantov: problémy sa snažíme riešiť hneď bez opakovania učiva. Čo sa stane: deti rozhodujú s učiteľom, ale riešenie nastáva na úrovni mechanického zapamätania, bez pochopenia. Preto, keď dostanú na skúške podobnú úlohu, nedokážu ju zvládnuť. Opakujem: pri riešení problémov nebolo pochopenie.
Prejdime k praxi.
Používame výber problémov z webovej stránky „Vyriešim jednotnú štátnu skúšku“ od Dmitrija Gushchina. Čo robí tento zdroj atraktívnym, je to, že prakticky neexistujú žiadne chyby a štandardy odpovedí sú dobre napísané.
Pozrime sa na problém C 6 č.12018.
Sada chromozómov somatických buniek pšenice má 28.
Určte chromozómovú sadu a počet molekúl DNA v jednej z buniek vajíčka pred začiatkom meiózy, v anafáze meiózy 1 a v anafáze meiózy 2. Vysvetlite, aké procesy prebiehajú v týchto obdobiach a ako ovplyvňujú zmenu počtu DNA a chromozómov.
Prvky odpovede:
Bunky vajíčka obsahujú diploidnú sadu chromozómov - 28 (2n2c).
Pred začiatkom meiózy - (2n4c) 28 xp, 56 DNA
V anafáze meiózy 1: (2n4c = n2c+n2c) - 28 xp, 56 DNA.
Meióza 2 zahŕňa 2 dcérske bunky s haploidnou sadou chromozómov (n2c) – 14 chromozómov, 28 DNA.
V anafáze meiózy 2: (2n2с= nc+nc) - 28 chromozómov, 28 DNA
Úloha je zložitá, ako pomôcť absolventovi pochopiť jej riešenie?
Jedna možnosť: nakreslíme fázy meiózy a ukážeme všetky manipulácie s chromozómami.
Akčný algoritmus:
- Pozorne si prečítajte problém, definujte úlohu, zapíšte si fázy, v ktorých potrebujete uviesť množstvo genetického materiálu
a) Pred začiatkom meiózy
b) V anafáze meiózy 1
c) V anafáze meiózy 2
- Vytvorte nákresy pre každú určenú fázu meiózy a vysvetlite, čo ste urobili.
Dovoľte mi objasniť: nepoužívame kresby, vyrábame ich sami. Táto operácia funguje na pochopenie ( aj keď strácame v estetike, nakoniec vyhráme!)
1. Pred začiatkom meiózy
Vysvetlím: meióze predchádza interfáza, v interfáze dochádza k zdvojeniu DNA, preto je počet chromozómov 2n, počet DNA je 4c.
2. V anafáze meiózy 1
Vysvetlím: v anafáze meiózy 1 sa chromozómy rozchádzajú k pólom, t.j. Z každého páru homológnych chromozómov sa do dcérskej bunky dostane len jeden. Sada chromozómov sa stáva haploidnou, ale každý chromozóm pozostáva z dvoch chromatidov. Keďže bunkové delenie ešte neprebehlo a všetky chromozómy sú v jednej bunke, chromozómový vzorec možno zapísať ako: 2n4c (n2c+n2c) 28 xp, 56 DNA (14 xp 28 DNA + 14 xp 28DNA)
3) V anafáze meiózy 2
Anafáza meiózy 2 nastáva po prvom (redukčnom) delení. Sada chromozómov v bunke p2c. Počas anafázy meiózy sa delia 2 centroméry spájajúce sesterské chromatidy a chromatidy sa, ako pri mitóze, stávajú nezávislými chromozómami. Počet chromozómov sa zvyšuje a rovná sa 2n2c. A opäť, keďže bunkové delenie ešte nenastalo a všetky chromozómy sú v jednej bunke, chromozómovú sadu možno zapísať nasledovne: 2n2c (nc + nc) 28 xp, 28 DNA (14 xp 14 DNA + 14 xp 14 DNA).
- Zapíšte si odpoveď. (máme to uvedené vyššie)
Aby som to zhrnul: Riešenie problémov tohto typu si nevyžaduje snahu o kvantitu, tu je dôležité dosiahnuť pochopenie logiky riešenia a poznanie správania sa chromozómov v každej fáze delenia.
Použité zdroje:
- FIPI “Metodické odporúčania k niektorým aspektom skvalitnenia výučby biológie”, autor. G.S. Kalinová, R.A. Petrošová. Moskva, 2014
- Biológia. Všeobecné vzory 10. ročník: učebnica pre vzdelávacie inštitúcie / V.B. Zakharov, S.G. Mamontov, N.I. Sonin - Moskva: Vydavateľstvo dropa, 2011.
- Budem riešiť Jednotnú štátnu skúšku. http://bio.reshuege.ru/
Bunková teória, jej hlavné ustanovenia, úloha pri formovaní moderného prírodovedného obrazu sveta. Rozvoj vedomostí o bunke. Bunková štruktúra organizmov, podobnosť štruktúry buniek všetkých organizmov je základom jednoty organického sveta, dôkazom príbuznosti živej prírody.
Bunka je jednotka štruktúry, životnej aktivity, rastu a vývoja organizmov. Rozmanitosť buniek. Porovnávacie charakteristiky buniek rastlín, živočíchov, baktérií, húb.
Štruktúra pro- a eukaryotických buniek. Vzťah medzi štruktúrou a funkciami častí a organel bunky je základom jej integrity. Metabolizmus: energetický a plastový metabolizmus, ich vzťah. Enzýmy, ich chemická podstata, úloha v metabolizme. Etapy energetického metabolizmu. Fermentácia a dýchanie. Fotosyntéza, jej význam, kozmická úloha. Fázy fotosyntézy. Svetlé a tmavé reakcie fotosyntézy, ich vzťah. Chemosyntéza.
Biosyntéza bielkovín a nukleových kyselín. Maticový charakter biosyntetických reakcií. Gény, genetický kód a jeho vlastnosti. Chromozómy, ich štruktúra (tvar a veľkosť) a funkcie. Počet chromozómov a ich druhová stálosť. Stanovenie súboru chromozómov v somatických a zárodočných bunkách. Životný cyklus bunky: interfáza a mitóza. Mitóza je delenie somatických buniek. meióza. Fázy mitózy a meiózy. Vývoj zárodočných buniek u rastlín a živočíchov. Podobnosti a rozdiely medzi mitózou a meiózou, ich význam. Bunkové delenie je základom pre rast, vývoj a rozmnožovanie organizmov.